
Biotechnológikaland
Ukrajna és a GMO – Dakotarege
(Géntechnologikaland No78) Ukrajnában a GMO-k társadalmi megítélése összességében óvatos, sokszor elutasító. A közvélemény-kutatások és piaci tapasztalatok szerint a lakosság jelentős...


(Biotechnológikaland No35)
Európában a növényi géntechnológia által képviselt legnagyobb buktató egy – idegenbeporzó növényeket érintő – biológiai alapú folyamat, ami valójában érdekvédelem által motivált gazdasági ok. Fajtahibrid-képződésnek nevezzük. A nemzeti fajták forgalmát veszélyeztetik a szabadalommal bíró nemzetközi tulajdonú GM-fajták (a globalizáció így kerül képbe), és ez az EU tagállami nemesítők számára nem igazán jó hír. Az egyidejű termesztéskor (vö. termesztési koegzisztencia) a keresztbeporzás az ökológiai termesztőket hozná kezelhetetlen helyzetbe, hiszen náluk kizárt a géntechnológiai úton módosított fajták termesztése, így ennek kereszteződésben való előfordulása is. Az EU-ban vetésre engedélyezett MON 810 kukorica a pollenjében lévő idegen génnel (cry1Ab) már az évben Cry1Ab-toxint termeltet a fajtahibridjeiben, így az eredetileg biotermesztésre szánt fajtánk szemeiben is, vagyis a keresztbeporzás révén felülírja az ökológiai termelés célját. A fajhibrid-képződés Európában főként a GM-repce engedélyezése esetén jelentene komoly gondot, ami keresztbeporzással a káposzta- (Brassica) és a retekféléket (Raphanus) érintené. Európában a cukorrépafélék ősei (Beta) gyomnövényként élnek, míg a kukorica teosinte nevű ősei (Zea), a hírek szerint, mint invazív gyomnövények éppen Spanyolországban terjednek, ahol a MON 810 GM-fajtacsoportot is termesztik.

A hibridképződés az olyan jelenségek közé tartozik, amelyek felkavarják a képzeletünket. Jellemző talán erre a ’Hibrid’ (Splice) című sci-fi horror, ami azzal az érzéssel is szembesített, hogy mennyi előítélettel gondolunk az ismeretlenre. A film alapján azt sem állíthatom, hogy ok nélkül. Az új fajok születése (speciáció) és kihalása (extinkció) az evolúció számára hétköznapi jelenségek. Ha a faj sikeresen alkalmazkodik az új környezetéhez, akkor megmarad (ilyenkor esetleg igénybe veszi a forrást a már korábban létezők elől), ha nem, akkor eltűnik. Környezetünkben az új fajok születése azon a szinten, amit érzékszerveinkkel nem érzékelünk (értsd mikrobiális élőlények), igen gyakori, de növényeket és rovarokat tekintve sem vesszük észre az orrunk előtt keletkező újakat. Összehasonlító/elemző képességünket messze meghaladja az a tudás, ami ehhez kellene. A természettudományi múzeumok szobáiban, gyűjteményi háttérrel van esély a meghatározásra. Persze létezik örökítőanyag-meghatározáson alapuló elkülönítés is, de ma még nem ez a jellemző a biológia által szemlélt látható világban.
A fajképződés lehetséges esete az is, hogy van két eltérő, ám rokon fajunk, amelyek eddig elkülönülten éltek, de most találkoztak/párosodnak, és így egy harmadik faj képződik. Ez esetleg hasonlíthat ahhoz az ősi fajhoz, amelyből valamikor elkülönültek. Lényeges, hogy az utódok életképesen örökítsék az új tulajdonságaikat. Létrejöhetnek ugyanis olyan élőlények is, amelyek kimérák, vagyis két faj/egyed szövetei egy egyedben, de örökíteni csak az egyik fajt/egyedet képesek, ha egyáltalán szaporodásképesek. A kiméra tehát egyedien új élőlény, de a speciáció számára zsákutca, amelyben az egyedi formának nincs folytatása. Van viszont olyan, ahol utód is születik, csakhogy az meddő. A leggyakrabban emlegetett példája ennek az öszvér, aminek szülői a ló és a szamár.
A kereszteződés azonban lehet a szaporodás szempontjából is sikeres. Ilyenkor természetes körülmények között a pioneer nemzedék megmérkőzik a táplálékforrásért és annak fogyasztóival abban a niche-ben, amelyben létrejött. Ezt tekintve az emberi evolúció is figyelmet érdemlő, hogy miként maradtunk a nemzettségünkben (Homo) társfaj nélkül. Ha az új minőség sikeres az alkalmazkodásban, akkor megveti a lábát és sokasodik, végül dominálhat is. A növényekre váltva, a rovarevő kancsóka-félék (Nepenthes) különösképpen ismertek arról, hogy számos természetes hibridfajuk létezik, amelyek a Fülöp-szigeteki és indonéziai területeken jöttek létre, és máig jól elkülöníthetően megmaradtak.
A termesztett növényeket illetően jól dokumentált példákat sorolhatunk. A szekunder hexaploid trikálé (100. kép) a hexaploid búza (Triticum) és a diploid rozs (Secale) keresztezésével, valamint oktaploid utódjaik és a primer hexaploid tritikálé (tetraploid búza x diploid rozs) visszakeresztezéssel jött létre. Ennek létrehozása már kifejezetten igényelte a nemesítők hatékony genetikai munkáját. Az ún. mutációs nemesítés eszközei (pl. besugárzás, EMS) elég változatosak, és eredményeik (pl. rizs, árpa, szója) nemzetközileg ismertek. Ugyancsak sikeresen alkalmazták a poliploidizáció jelenségét több termesztett növényünk (pl. búza, kivi, görögdinnye) esetében, amelyeket nem követett olyan mértékű közfigyelem, mint a növényi géntechnológiát.

100.kép: Tritikálé (Fotó: camli.com)
A géntechnológia egyik mai célpontja az olajrepce (Brassica napus) is fajhibrid (5. ábra – lásd az No7 részben), amennyiben a tarlórépa (Brassica rapa) és a káposzta (Brassica oleracea) utódja. A Brassica-fajok géncentruma a Mediterráneumban található, vagyis itt találjuk a legtöbb közeli rokonfajt, amelyekkel sikeres kereszteződés valósulhat meg. Különösképpen a hímsteril Brassica napus (amelyeket éppen a hibridképződés megakadályozására hoztak létre ebben a GM-fajtacsoportokban) beporzódhat a repcsénretektől (Raphanus raphanistrum), de a termesztett retektől (Raphanus sativus) is. A közelrokon fajok között tehát természetes úton is létrejöhetnek interspecifikus hibridek (101. kép). Nem jellemző ez a távoli fajokra, így a bakteriális eredetű transzgénekre, pl. a Bt-növényekbe géntechnológiai úton vitt cry-génekre.

101.kép: A retek (Raphanus sativus) és a kínai brokkoli (Brassica alboglabra) hibridje (Fotó: Li, X. et al.)
A növényekre az önbeporzás és az idegenbeporzás a jellemző. Az önbeporzókból a fajidegen gén tovább kerülése alacsony valószínűségű. Ezeknél a keresztbeporzást néhányszor tízméteres izolációs távolság is megakadályozza, vagyis valamilyen közvetítő szervezet (pl. rovarok) kell a hibridek létrehozásához. Más a helyzet az idegenbeporzóknál, ahol a szél- vagy vízbeporzás, továbbá a rovarbeporzás (elsősorban méh-, légy- és lepkefélék), de madár- és denevérfajok is általi beporzás is ismert (102. kép). A transzgén terjedésének megakadályozásához itt az izolációs távolságnak sokkal nagyobbnak kell lennie.
A 30. ábrán feltüntetett izolációs átlagadatok még a GM-fajtacsoportok termesztési gyakorlatban való megjelenése előtti értékek, amikor a fajtahibrid-képződés még komolyabb (értsd jogi rendezés) problémával nem járhatott. Az ábra adatainak értékeléséhez tudnunk kell, hogy az önbeporzóknál is előfordulhat egyidejű rovarbeporzás, esetleg szélbeporzás is. A megadott izolációs távolságok általában a minimális – termesztői tapasztalatokra alapozott – értéket rögzítik, ami nem jelent mentességet; csupán azt, hogy az állományunk gazdasági szempontból egyöntetű marad. A rovarbeporzású növényeknél sokan csak a mézelő méhre gondolnak, de ez koránt sincs így. Tény, hogy a mézelő méh közelítőleg három kilométeres sugarú körben gyűjt, így a megporzási körzete ezzel jellemezhető. A szélbeporzásról is jó, ha tudjuk, hogy az egyes növények pollenjeinek morfológiája eltérő lebegőképességet biztosít a számukra. A nagyobb gömb alakú pollen (pl. kukorica) viszonylag gyorsan kiülepedik, míg a nagy felületű, röpülést segítő képletekkel rendelkezők igen messze juthatnak. A pollenszórástól eltelt időtartam azonban nagyon kritikus, hiszen a pollen a kiszáradásra és az UV-sugárzásra is igen érzékeny, vagyis életképessége – ami megtermékenyítő képességével jellemezhető – rövid. Sok faj (pl. kender) pollenje messze ellebeghet ugyan, de időközben a megtermékenyítő képességét elveszítheti.
A fajtatiszta vetőmag előállításához szükséges ún. izolációs távolságot a különböző szaktekintélyek sokszor eltérőnek gondolják. A kukorica-vetőmag termesztésre hazánkban 400 méteres izolációs távolság vonatkozik (lásd később), azonban az Egyesült Államokban a csemegekukorica és a pattogató kukorica előállításához 1600 méteres izolációs távolságot javasolnak. Az általános igazságot, mégpedig, hogy a nagy kukoricapollen rövidtávon könnyen kiülepedik, felülírhatja a pollenszóráskor uralkodó szeles időjárás, amely ilyen pollencsóvát is felemelhet, majd nagyobb távolság után kiszór. Nem is véletlenek a kukoricával kapcsolatos vetőmag-tisztasági problémák, amit hazánkban kiszántások követtek.
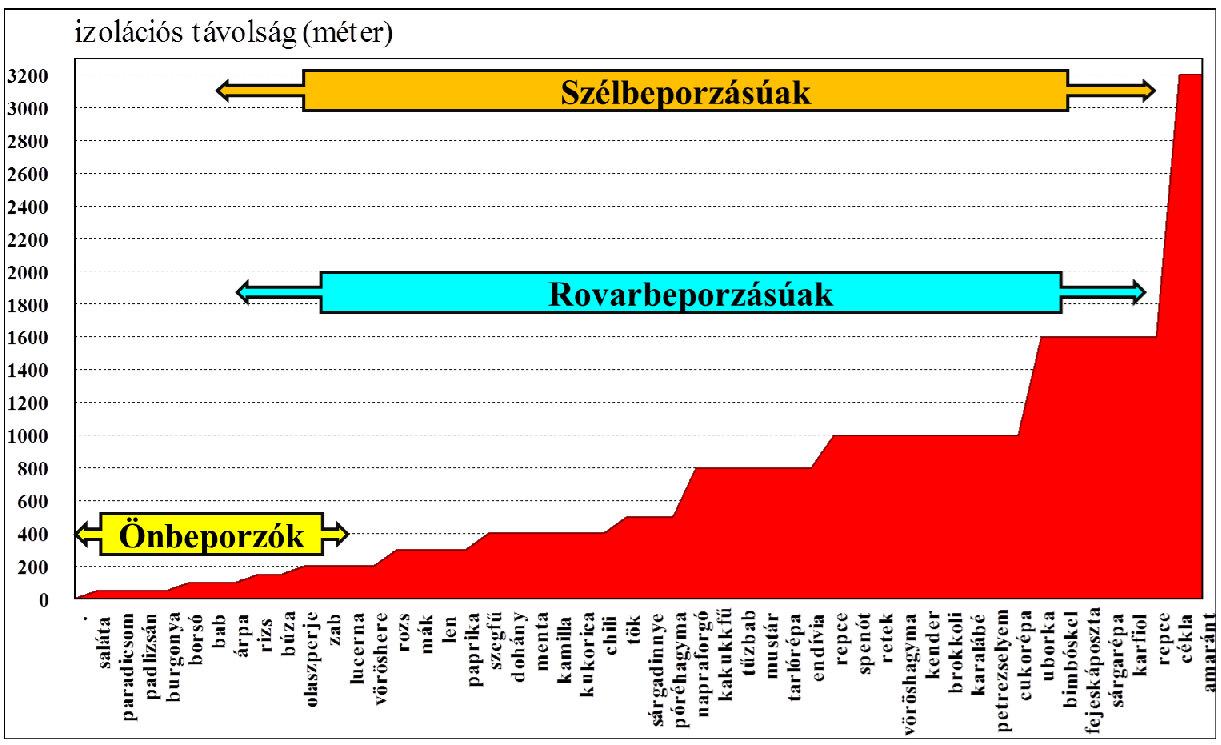
30.ábra: Vetőmag-termesztésnél hazánkban és nemzetközileg javasolt izolációs távolságok
A fajhibridképződés valószínűségénél a fajtahibrid-képződésé (intraspecifikus hibridek) magasabb. Itt arról van szó, hogy az egyik fajtát egy másik fajta pollenje porozza be, így az utódnövények genetikai tartalma megváltozik. Ez kedvezőtlenül hat az ún. fajtatisztaságra. A megjegyzésem a szabadon elvirágzó fajtákra vonatkozik, amelyekből saját magunk vetőmagot foghatunk. Az ún. hibridfajtáknál más a helyzet. Itt általában két beltenyésztett vonal utódmagjait vásároljuk vetőmagként, s ez a generáció a hibridvigor jelensége miatt magas teljesítőképességű. Viszont ez a tulajdonság a következő generációban elvész (hasad a genetikai tartalom), vagyis saját vetőmag fogása nem javasolt. Ehhez képest a GM-fajtáknál tilalmas is – szabadalom védi ezt a lehetőséget – vagyis a fajta szabadalmi tulajdonosának minden évben meg kell fizetni az ún. technológia díjat.
Mielőtt tovább haladunk, vessünk egy pillantást a faj (species) és a faj alatti taxonómiai egységekre. Mikroorganizmusoknál már a fajok elkülönítésekor is zavarba jöhetünk, de soksejtűeknél talán már nem. A klasszikus morfológia az élőlény kinézete alapján különít el. Növényeknél ebben fontos szerepet játszik a virág szerkezete. A faj alatti kategóriák alfaj (subspecies), változat (varietas), alak (forma) már sokkal kevésbé könnyen körülírható fogalmak. A különböző szaktekintélyek szerint azt találhatjuk, hogy az egyik fajként, míg a másik alfajként (vö. Beta és Zea) tekint ugyanarra az élőlényre.

102.kép: Hemaris thysbe a selyemkóró (Asclepias syriaca) virágját porozza be (Fotó: Mary Holland)
A már említett kínai brokkolit egyesek önálló fajként tartják számon és Brassica alboglabra L.H. Bailey néven szólítják, addig mások Brassica oleracea var. alboglabra (L.H. Bailey) Musil néven egy faj változataként (cultivar.) kezelik. A Brassica oleracea fajon belül csoportok, változatok és formák sokaságát találjuk, amelyek végén különítjük el a fajtákat, amelyekre az egyformaságuk a leginkább jellemző. A fajtabejegyzés és -fenntartás ezt az egyöntetűséget védi (hazánkban is fajtapénz jár érte a nemesítőknek, vagyis nyilvántartott és honorált teljesítmény), és a fajtatisztaság kifejezést használjuk a védeni kívánt alapállapotra, amelyben idegen gének megjelenése nem kívánatos. Ez a növénynemesítők ellenkezésének alapja lehet, hiszen a transzgének kimutatása nem bonyolult feladat, és mivel természetes körülmények között nem fordulnak elő, ezért bizonyításra használhatók.
A fajtahibrid-képződés idegenbeporzó fajoknál gyakori esemény. A GM-fajtáknál azért emelkedik ez különlegesen fontos tényezővé, mert megváltoztathatja az eredeti termelési szándékot. Az Európában vetésre engedélyezett MON 810-es kukorica például cry1Ab-gén örökít a pollennel. Ha ilyen fajta termesztője biokukorica-termesztő szomszédságába települ és a termesztett fajták virágzása egyidejű, akkor fajtahibridek képződhetnek mindkét táblán. Akkor is, ha betartják a hivatalosan előírt 400 méteres izolációs távolságot. A GM-táblán keletkező fajtahibrid-magok nem okoznak problémát, mert vetőmagnak – a GM-fajtatulajdonossal kötött szerződés szerint – eleve nem alkalmazhatók. Az ökológiai termesztés területén viszont kisebbségben ugyan, de olyan magok is érlelődnek, amelyhez a pollen a MON 810-es kukoricától származott, és az új magban ez a gén Cry1Ab-toxin termelésébe kezd. A keresztbeporzás itt azonnal törli az ökológiai termesztő eredeti elképzelését, hiszen a betakarított terméke (áru- és vetőmag-kukoricája) már nem lesz biominőségű, ahogy azt a termesztési szabályai a számára előírták, mivel olyan bakteriális toxint tartalmaz, ami természetes körülmények között kukoricában nem fordulhatna elő. Hangsúlyos, hogy ezt most nem toxikológiai megfontolás utáni véleményként írom, hanem egy önálló termesztési ágazat rendjébe való avatkozás eseteként értékelem, amit a hazai jog véd. Ki követi el azonban a hibát – ki fizet kártérítést? –, ha az előírt izolációs távolság elégtelen?
Megdöbbentő, hogy ilyen esetben a GM-fajta szabadalmasa – ha bizonyítja idegen táblán a szabadalmaztatott génjének megjelenését – perelhet a technológia díj elmulasztása miatt, holott a beporzást esetleg a szél vagy a rovarok intézték el. Az ebbéli félelem a lényegi gazdasági oka a biotermesztőkön túlmenően az intenzív módon gazdálkodó, módosítatlan fajtákat termesztők növény-géntechnológia ellenzésének. Egy környezetünkbe kibocsátott, szaporodóképes szervezet szabadalmi tulajdonként való kezelése talán mégsem volt jó ötlet. A biológiai törvényei jogi eszközökkel nem írhatók felül.
A génáramlás/génsodródás jelenségei a növényi géntechnológia esetében a kézben tarthatóságot megkérdőjelezik, míg a zárt tartású GM-állatok esetében kevesebb ellenkezést válthatnak ki. A termelői és fogyasztói reakciókat majd csak ezt követően látjuk.
Az azonos és rokon fajok közötti ivaros géncserén (vertikális géntranszfer) kívül bonyolultabb mechanizmusok (transzdukció, transzformáció, konjugáció, transzpozon-aktivitás) is léteznek, amelyek során igen távoli élőlények génjei átkerülhetnek magasabb rendű élőlényekbe. Az ún. horizontális géntranszfer során tehát távoli fajok közötti speciális géncsere is megvalósulhat. Erre legismertebb példa az antibiotikum-rezisztencia génjeinek – plazmidokon foglalnak helyet – átadása a mikroorganizmusok között, de a vírusok közé tartozó bakteriofágok is átvihetnek a forrástól eltérő fajba DNS-darabokat.
A xenotranszplantációs kutatások során figyeltek fel a sertés genetikai állományában található endogén retrovírusokra, amelyekhez hasonlók az ember esetében is felismerhetők. Egy felmérés szerint az emberi 20 ezer gén közül közel százötven idegen eredetű. Úgy gondolható, hogy az ebbéli kívánatos tudásunknak még csak a kapujában állunk. Sokan erre alapozva az emberi géntechnológiai tevékenységet a horizontális géncseréhez sorolják, és természetesnek gondolják. Ez esetben a genetikai információ bővülésének lehetőségét a géntechnológia gyakorlati művelőinek genetikai tudása – annak korlátaival együtt – teremtette meg. Illene ezen a tudásterületen csak biztosra menni. Az európai szkepszis azonban meglehetősen nagy, mert a biztosítékok a pillanatnyi tudásunkra vonatkoznak.
Térjünk vissza a növényi géntechnológia gyakorlatához, és gondoljuk végig a kukorica példáját. Itt mindjárt két termelési célt külön kell választanunk. Az árutermesztés pollen-kompetitív viszonyok között történik, vagyis a közelben lévő egyedek megporzási esélyei sokkal jobbak, mint a távoli pollenforrásé. A vetőmag-előállításnál viszont eltávolítjuk a hímvirágokat (kukoricánál címerezünk) vagy hímsteril anyavonallal (repcénél gyakori) dolgozunk. Itt nincs pollen-kompetíció, vagyis megnő az esélye a távoli pollenforrásoknak. Bálint Andor vetőmag-termesztésről írt könyvében az olvashatjuk (31. ábra), hogy árutermelésnél, 500 méternél 0,5%, míg vetőmag-termesztésnél 2,6% a mérhető hibridképződési arány. Ez utóbbi – vetőmagtermesztők figyelem! – azt jelentené GM-fajtaszomszédságnál, hogy az egyébként módosítatlan fajtánkat GMO-ként kell jelölnünk.
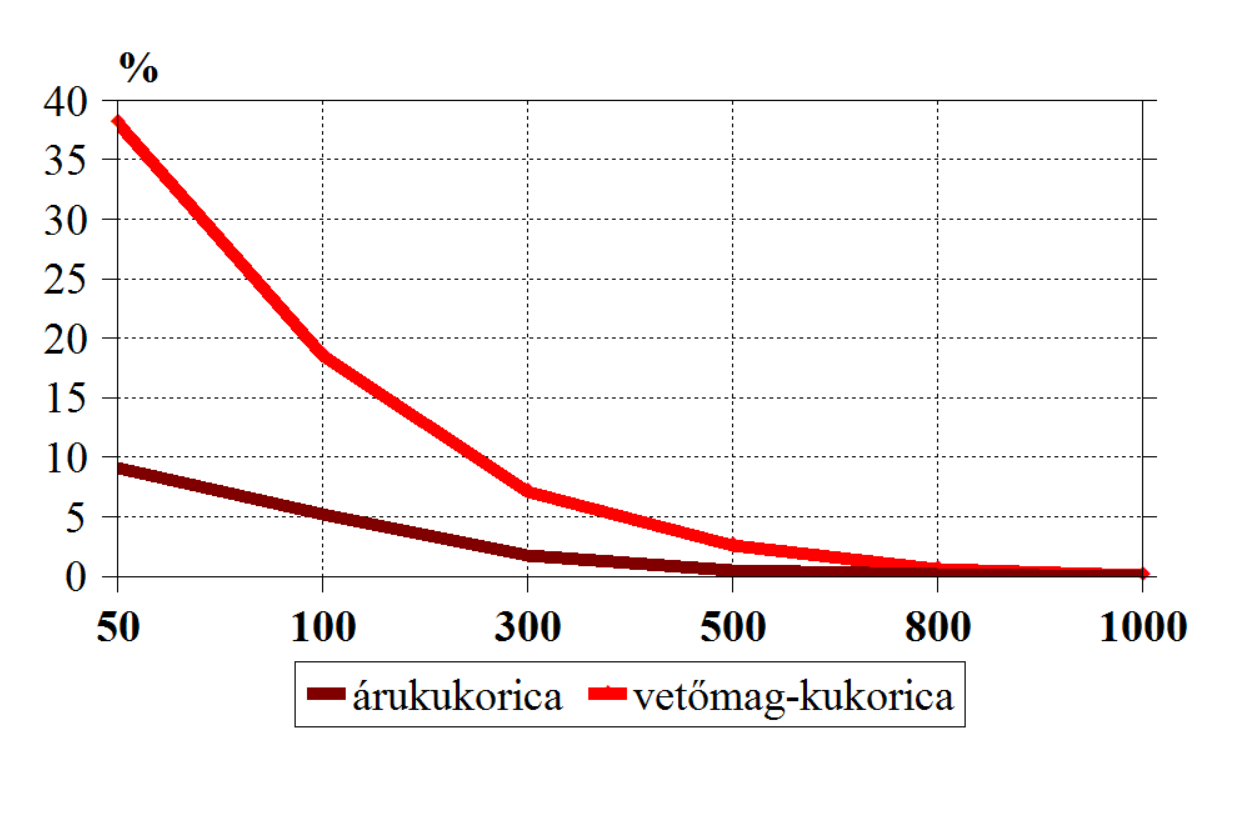
31.ábra: A fajtahibrid-képződés valószínűsége fehér kukoricánál az idegen (sárga) pollenforrástól távolodva (Forrás: Bálint Andor, 1980)
800 méternél 0,6% hibridképződést mértek vetőmag-kukoricánál. Ez már nem jelölésköteles szennyezés ugyan, de nem megfelelő minőségű vetőmagot jelent, aminek hatása hosszú távon nem elfogadható. Az általunk végzett kísérletben 800 méternél már nem találtunk hibridképződést, vagyis a csövek csupaszok maradtak, azonban mi kis kiterjedésű kék (virágzása kicsit koraibb volt) és sárga (virágzása teljesen egybe esett) pollenforrásokkal és sárga címerezett anyavonallal dolgoztunk (103. kép). A fajtahibrid-képződés mellett további problémát jelent az országba beszállított vetőmagvak kérdéses tisztasága, a vetőmagvak kezelésének zavarai, a fizikai keveredés is.

103.kép: Címerezett sárga kukoricán az uralkodó szélirányban termékenyülő szemek eltérő távolságú és kissé eltérő virágzási idejű sárga és kék pollenforrásnál (Fotó: Darvas Béla)
A fentiek szerint, míg az árukukoricánál (kivéve biokukoricát) elégséges lehet a ma is jogszabály szerint érvényben lévő 400 méteres izolációs távolság, sőt a távolság csökkenthető lehetne, ha hatsoros szegélysort vetünk a GM-tábla köré, ami a pollenkilépést megakadályozza, addig vetőmag-termesztésnél lényegesen hosszabb izolációs távolságot kellene biztosítani. Mindez azonban nem abszolút biztosíték. Vizsgálataink során tapasztaltuk, hogy 800 méterre az uralkodó széliránnyal szemben a tábla közepén is előfordult pár megporzott kék színű szem, amit kukoricabogarak közvetítettek. A kukoricabogár (Diabrotica) nem ismert, mint pollinátor, viszont érési táplálkozásában a kukorica címere fontos szerepet játszik, ami után a kukorica bibéin táplálkozik. E közben helyváltoztatással a testén hurcolt pollennel meg is termékenyíthet. Ez a jelenség az átlagra nem jellemző ugyan, de megtörténik.
Hosszabb időtávon az ilyen módon keletkező szemekből a transzgének terjedése már kikerül az ellenőrzésünk alól. Így számomra érthető, ha Heszky László (növénygenetikus) és †Balla László (növénynemesítő) arra a következtetésre jutottak, hogy egy országnak a kezdetekkor el kell dönteni, hogy egy adott kultúrában, idegen beporzó fajoknál GM-fajtákat hasznosít vagy sem. Ez egyidejűleg – esetleg – az adott faj hazai nemesítésének további szükségességét is megkérdőjelezné.
Összegezve a faj- és fajtahibridek kérdését: az előző ritkább és speciálisabb eseményeket feltételez, továbbá a gyakorlatban való előfordulásuk a GM-repce európai engedélyezése után a Brassica és Raphanus fajok esetében valószínűsíthető. A cukorrépát európai gyomfajból (sziléziai fehérrépa) nemesítették, míg a teosinte-fajok (kukoricával rokon gyomok – 104. kép) – amelyek eredetileg Közép-Amerikában élnek – Spanyolországban és Franciaországban éppen terjedőben lévő gyomnövények, s a MON 810 kukorica termesztésével hozzák ezt a történetet összefüggésbe. Ez esetben a transzgén új fajba kerülve kikerülhet az ellenőrzésünk alól, leginkább, ha a gyomflóra részévé válik (vö. repcsényretek x GM-repce, teosinte x GM-kukorica). Ehhez képest a fajtahibrid-képződés idegenbeporzóknál természetes jelenségként fordul elő, és eddig szabályozási szempontból kezelhetetlen problémának minősíthető Európában.

104.kép: A teosinte egyik faja: Zea diploperennis (Fotó: teosinte.wisc.edu)
A következő rész címe: Hatás kontra ellenhatás – Rezisztencia (Biotechnológikaland No36)
Darvas Béla
Bankszámlaszám: 12011265-01425189-00100001
Bank neve: Raiffeisen Bank
Számlatulajdonos: Átlátszónet Alapítvány
1084 Budapest, Déri Miksa utca 10.
IBAN (EUR): HU36120112650142518900400002
IBAN (USD): HU36120112650142518900500009
SWIFT: UBRTHUHB
Számlatulajdonos: Átlátszónet Alapítvány
1084 Budapest, Déri Miksa utca 10.
Bank neve és címe: Raiffeisen Bank
(H-1133 Budapest, Váci út 116-118.)
Támogasd a munkánkat az Átlátszónet Alapítványnak küldött PayPal-adománnyal! Köszönjük.
Támogatom PayPal-adománnyalHa az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
1% TÁMOGATÁS


Támogasd a munkánkat 10 ezer forint adománnyal, mi pedig megajándékozunk egy pólóval. Katt a részletekért.
Támogasd a munkánkat palackvisszaváltással, kattints az üvegvisszaváltós oldalra, mentsd el a kódunkat, és használd azt a Repontoknál!
Ha van bankkártyád, akkor pár kattintással gyorsan tudsz rendszeres vagy egyszeri támogatást beállítani nekünk az adjukossze.hu oldalán.


Postai befizetéssel is tudsz minket támogatni, amihez „sárga csekket” küldünk. Add meg a postacímedet, és már repül is a csekk.
Néző, Szurkoló, B-közép és VIP-páholy kategóriás Átlátszó-előfizetések között válogathatsz a Patreonon.
Bárhol is dolgozol a világban, ha a munkáltatód lehetőséget ad arra, hogy adott összeget felajánlj egy nonprofit szervezetnek, akkor ne feledd, a Benevity-n keresztül az Átlátszónet Alapítvány is ajánlható.
Ha az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
(Géntechnologikaland No78) Ukrajnában a GMO-k társadalmi megítélése összességében óvatos, sokszor elutasító. A közvélemény-kutatások és piaci tapasztalatok szerint a lakosság jelentős...

(Biotechnologikaland No77) A Marton Genetics Kft. állásfoglalása A Marton Genetics (Bázismag Kft.) cégcsoport a martonvásári kutatóintézet (Agrártudományi Kutatóközpont) munkájára épült....
(Biotechnologikaland No76) Előszó A GMO-lobbi hatására az Európai Bizottság javaslata (címábra) komoly visszhangra talált Európában. A javaslatok közül NGT1 esetében...

(Biotechnologikaland No75) Barabás Zoltán Biotechnológiai Egyesület kontra GMO-Kerekasztal A Barabás Zoltán Biotechnológiai Egyesület lépett hazánkban először abból a célból, hogy...
Támogasd a munkánkat banki átutalással. Az adományokat az Átlátszónet Alapítvány számlájára utalhatod. Az utalás közleményébe írd: „Adomány”, köszönjük!