
Biotechnológikaland
Ukrajna és a GMO – Dakotarege
(Géntechnologikaland No78) Ukrajnában a GMO-k társadalmi megítélése összességében óvatos, sokszor elutasító. A közvélemény-kutatások és piaci tapasztalatok szerint a lakosság jelentős...

(Biotechnológikaland No31)
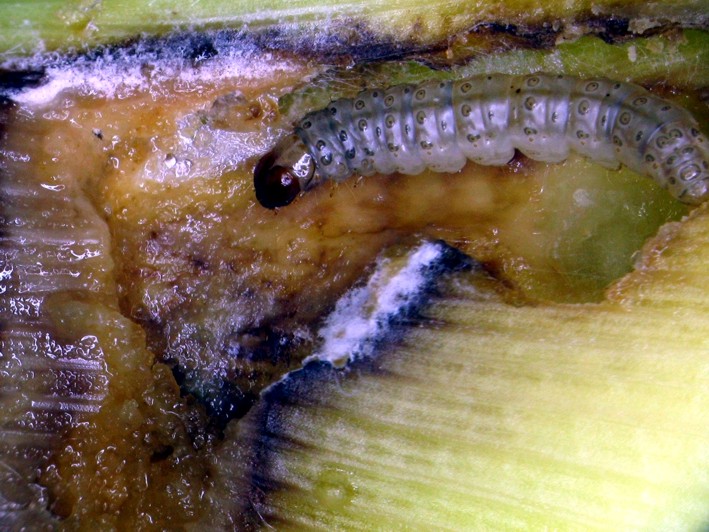
A MON 810 kukorica-fajtacsoport vetési moratóriumával nagyon szerteágazó kísérleteket végzett az a kutatócsoport, aminek tagja vagyok. Az eredményeinket sokan interpretálták, túlozták el azokat pro vagy kontra. A sorozat következő fejezeteiben ezeket veszem sorra. Első ezek közül a Bt-kukorica által a területegységen megtermelt Cry-toxin mennyisége, ami a fehérjetoxin permetező szer formájában kijuttatott változatához képest 50-1500-szoros mennyiségű. A Bt-növények ökológiai gazdálkodásából való kizárását már az is indokolhatná, hogy ezek a növények a természetes protoxinváltozat kurtított/preaktivált változatát termelik. A folyamatos és nagy mennyiségű termelés még az integrált növényvédelem alapelveivel is ütközik, amennyiben megelőző jellegű és nem okszerű. A gyakorlatban lévő Bt-növények nem szövetspecifikusan módosított formák, vagyis a Cry-toxintermelés minden részüket érinti. Ott is termelnek, ahol a célkártevő meg sem jelenik.

A fajtatulajdonosok európai vetésre ajánlott első növényi fajtacsoportja a MON 810 genetikai esemény volt. A genetikai esemény (event) kifejezés a biológiai egyediségére utal, amely egy kísérlet során jött létre, s amelynek utódnövényeiből fejlesztik ki a szóban forgó fajtákat. Az eseménynév (event name) első betűtagja az eredeti tulajdonosra utal. Többnyire a BCS – Bayer CropScience, a BPS – BASF Plant Science, DAS – a Dow AgroSciences, a DP – DuPont (Pioneer Hi-Bred International), MON – Monsanto, míg a SYN – Syngenta Seeds, fajtákat jelöli. A követő számkombináció az egyedi genetikai eseményhez rendelt azonosító. Akadnak eltérések – különösen a korai transzgenikus genetikai eseményekhez köthetően –, hiszen vannak vásárolt események, vagy felvásárolt cégek (pl. ACS – Aventis CropScience → BCS – Bayer CropScience), ahol az eseménynév változatlan maradt. Az eseménynévhez képest az eseménykód (ID-number vagy event code), ennek egyfajta tisztázata, ahol azért a számkombináció többnyire jól felismerhető. Ebből aztán számtalan kevert írásmód származott mára, ahol a helyközök használata, a Ø megjelenése a számkódban, a kiegészítő szám, valamint a kötőjeles, vagy nélküli írásmód keveredik.
Az első generációs – ma gyakorlatba került, többnyire növényvédelmi célú – különböző géntechnológiai módszerekkel (Agrobacterium, génpuska stb.) előállított fajták módosítatlan fajtákkal való keresztezésével jön létre a fajtacsoport, amelynek összefoglaló kereskedelmi neve (trademark – védjegy) is lehetséges. Például a glyphosate-tűrő növények egy jelentős csoportja Roundup ReadyTM névre hallgat. A konkrét fajtának azonban – ahol már egyedi nevek vannak – a közel izogenikus vonal fajtatulajdonosának vetőmag cége ad nevet. Például a hazánkba először kísérleti célra vetett MON 810 fajtacsoport (ezt hívják YieldGardTM, MaizeGardTM fajtacsoportnak is) a DeKalb Genetics Corporation (Monsanto → Bayer) vetőmag-céghez tartozó DK-440 BTY volt, ami arra utalt, hogy a közel izogénes anyavonala a DK-440. Valahogy a géntechnológia világában semmi sem egyszerű, és bizony képességpróbáló a pontos megértés. Már az elnevezésnél el lehet tévedni, és ez még nem is mezőgazdasági géntechnológia/genetika, amibe itt bele sem bonyolódunk, arra ott vannak a tankönyvek. Mivel a módosított apavonal és a módosított fajták genetikai összetétele közel sem azonos, ezért komoly tere nyílik a fajta képességeiben az összetétel jelentős szórására. Ez vonatkozik a Cry-toxinok termelésére is (később részletesen).
Maradjunk annyiban, hogy – többnyire a nemzetközi cégek laboratóriumaiban – a növényi géntechnológusok létrehoznak egy új, addig nem létező genetikai tartalmú módosított vonalat majd annak utódaiból egy fajtacsoportot és azt szabadalmaztatják. Ekkor olyan jogokat kapnak, ami a GM-vetőmag teljes körű felügyeletét hozzájuk rendeli. Megtilthatják és felügyelhetik a harmadik fél vizsgálatait. A vásárló csak a vetőmag használati jogát (technológiahasználati díjat fizet ezért) veszi meg, de nem rendelkezik szabadon vele. Nem adhat belőle harmadik félnek vizsgálatra, és nem foghat belőle magot (már a hibridfajtáknál sem tanácsos ez, de ott nem perelhető), nem használhatja további keresztezésekhez. Ennek megsértésekor a fajtatulajdonosok a jogaikat peres úton érvényesíthetik. Ez az a pont, ahol sokak gazdasági érdeke sérült (a gazdáé is, mert a magot csak lizingeli, de nem teljeskörűen birtokolja), ezért ennek az innovációnak az európai útja fölöttébb döcögős. Sok agrárpolitikus szerint a vetőmag stratégiai termék, aki birtokolja, az tartja kézben az érintett nemzet növénytermesztését. Az európai agrárpolitikusi fanyalgás lényege talán ez a félelem.
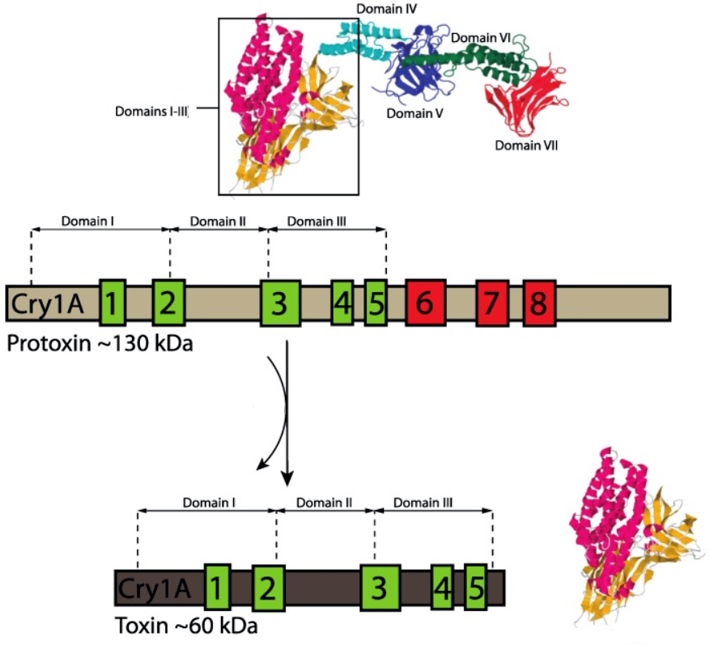
23.ábra: A Cry1Ab-protoxin aktiválódása három alegységű toxikus fehérjévé (Palma és mtsi nyomán)
Térjünk vissza a MON 810 fajtacsoporthoz, amely Európában először vetési engedélyt kapott, de amivel kapcsolatban főként Ausztria kezdeményezésére kifejezett tiltakozás indult el Európában. Ausztria civilszervezetei a velük szomszédos országokat is megszólították, mert egy módosított növények nélküli régiót akartak kialakítani. Tény, hogy mindez sikeressé vált, bár szerintem a kezdetekben nem sokan fogadtak volna hazánkban erre a fejleményre. A magyarországi MON 810 fajtacsoport vetési moratóriumának néhány környezettudományi eredményekre alapuló pontját mesélem itt el, amely vizsgálatokban személyesen is közreműködtem. Nem a vizsgálatokat fogom itt leírni, arra tudományos szaklapok és könyvek valók, hanem a vizsgálatok koncepcióit és az eredményekből levonható következtetéseket. Fontos az olvasónak megjegyezni, hogy az érvelésem mindig a konkrét esetre érvényes és nem általánosítható a módosított növények egészére, még ha némi áthallás van ebben az esetben más Bt-növényekre vonatkozóan is.
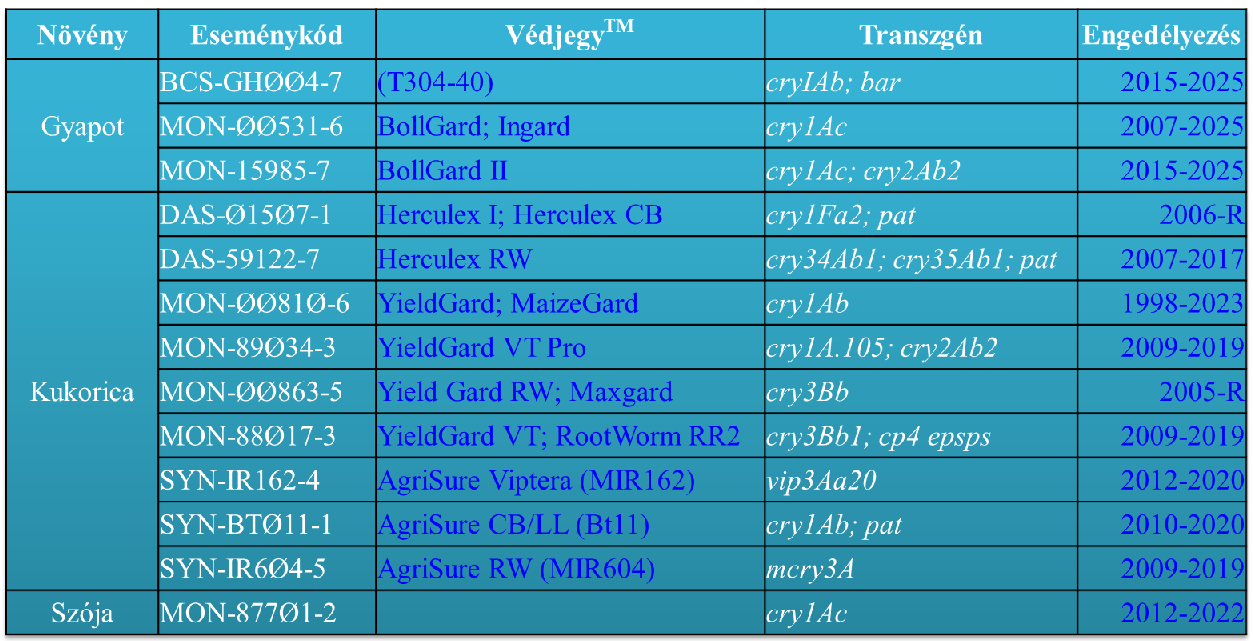
3.táblázat: Az Európai Unióban 2016-ban jelentős előrehaladást elért Bt-fajtacsoportok (R – felülvizsgálata folyamatban van)
A Bt-növények (3. táblázat) a Bacillus thuringiensis rovarpatogén baktériumnak a kurtított Cry-toxinjait termelik, bár már elérhető Vip-toxintermelő fajtacsoport is. A spóraképző baktériumban a fehérjetoxin kristályos formában (δ-endotoxin vagy parasporális test) van jelen (42-46 old.). A rovarok középbelében a kristályos szerkezet feloldódik és kiszabadul a protoxin, amit proteázok (pl. tripszin) hasogatnak (távolítják el a IV-VII alegységet/domént) és ezzel patogenitás szempontjából aktiválják azt. Így keletkezik az I-III doménű aktivált toxin (23. ábra). Ez a monomer aktivált toxinforma, ami a rovarbél különböző receptoraihoz (ALP – alkalikus foszfatáz, APN – amino-peptidáz és CAD – cadherin) kötődik és hat lépés után az oligomerjük a rovarbél falán pórust nyit (24. ábra), amin a B. thuringiensis vegetatív teste a lárva testüregébe kerülhet, ami annak halálához vezető szepszist idézi elő. A védekező szerként való alkalmazás során rájöttek arra, hogy élő B. thuringiensis baktériumra nincs is szükség a hatáshoz, mert a rovarbélben élő baktériumokkal ez a hatás ugyanúgy elérhető. A mai baktériumos készítményekben általában nincs is élő baktérium, sőt kizárólag csak Cry-toxin tartalmú permetező szerek is ismertek.
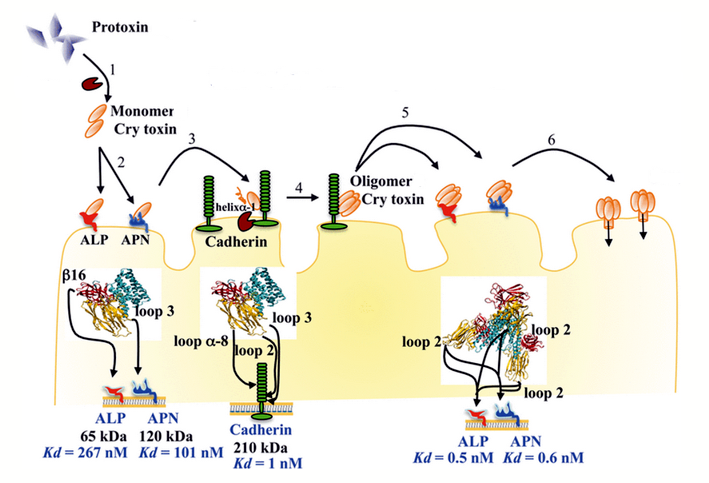
24.ábra: A Cry-toxin hatásmechanizmusa a rovar belében (Pardo-López és mtsai nyomán)
Mennyiben azonosak a különféle szerotípusok/törzsek által termelt Cry-toxinok a módosított Bt-növények által termelt kurtított (ún. preaktivált) toxinnal? Itt először a B. thuringiensis különböző szerotípusai által termelt fehérjetoxinok eltéréseire kell felfigyelnünk, ami szelektivitás-változással jár együtt. Például a B. thuringiensis serovar. aizawai, entomocidus, kurstaki és thuringiensis Cry1-, a serovar. kurstaki, kumamotoensis Cry2-, a serovar. morrisoni, sandiego, tenebrionis Cry3-, a serovar. israelensis Cry4-toxinokat termelnek, amelyek tömege/szerkezete jelentősen eltérhet (25. ábra). A különböző szerkezetű Cry-toxinok eltérő biológiai hatásúak és általában rendszintű specifitással rendelkeznek, de kivételek is vannak (26. ábra). A protoxin és az aktivált toxin közötti lényeges méretbeli különbség nem teszi lehetővé az egyszerű összefüggés feltételezését. A 131 kDa (1155 aminosav) tömegű Cry1Ab-protoxinnak például csak 60-65 kDa (594 aminosav) tömegű az aktivált, három alegységes formája. A MON 810-es kukorica ugyanakkor 91 kDa (816 aminosav) tömegű fehérjetoxint termel (ehhez a cry1Ab-gént kurtították), amit preaktiváltnak nevezünk.
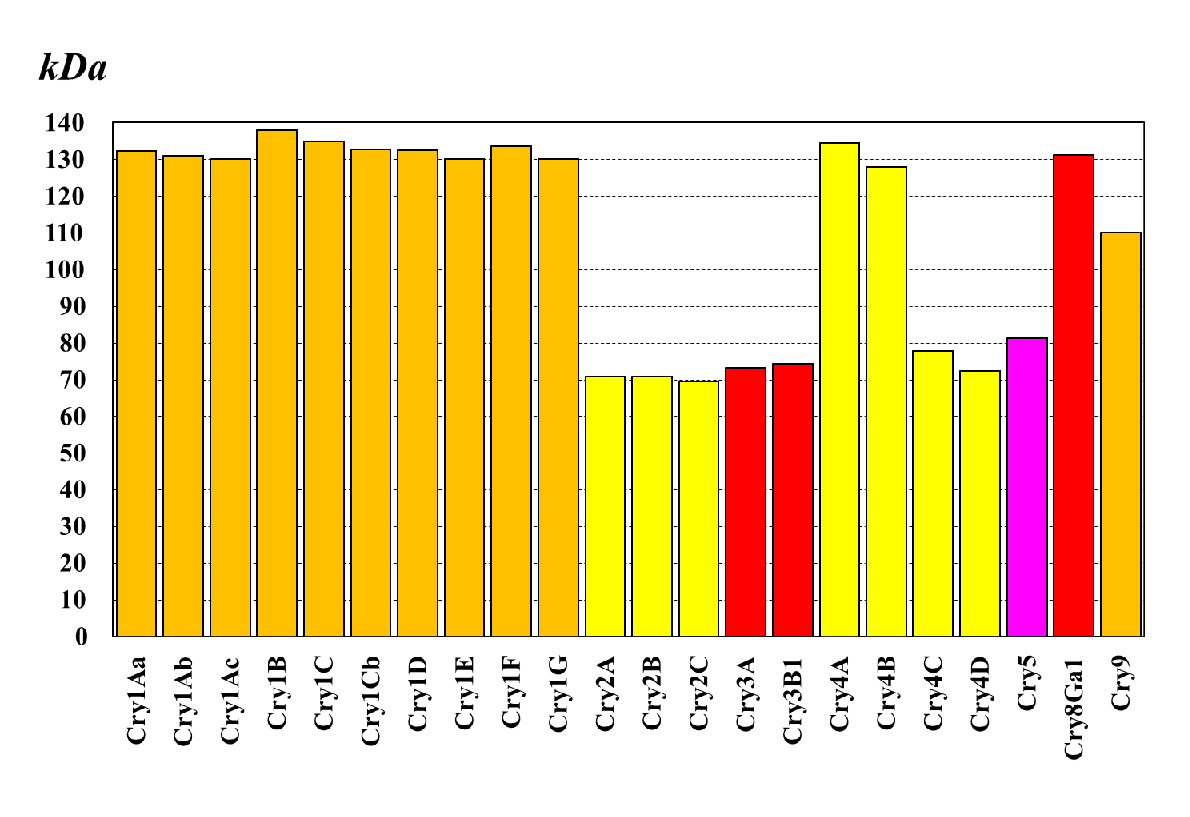
25.ábra: Különféle Cry-protoxinok molekulatömege (narancssárga – Lepidoptera; citromsárga – Diptera; piros – Coleoptera; lila – Nematoda fajokra specifikus hatásúak elsősorban)
Hogy ennél is tovább növeljük a bizonytalanságot, az egyaránt Cry1Ab-toxintermelőnek feltüntetett MON 810 91kDa (816 aminosav), az Európai Unióban már érvényes dossziéval nem rendelkező SYN-EV176 72,6 kDa (648 aminosav), a SYN-BT11 65 kDa molekulatömegű preaktivált toxinfehérjét termel. Helyes ezt így együttesen kezelni?
Bizonyosak lehetünk-e abban, hogy a Cry-toxinok nem lépik át az ismert szelektivitásuk határait? A fajtatulajdonosok ezt állítják, de néhány mérési eredmény bizonytalanná tehet bennünket (26. ábra). Az egyes toxinosztályokba tartozó alosztályok/csoportok hatását nagyon eltérő állatcsoportokban is ki tudták mutatni. A Cry-toxinok módosítása – amivel elérhető a rá lassabban kialakuló rezisztens rovarnépességek kialakulása – napjainkban újra az érdeklődés előterébe került. Jellemző példája ennek a B. thuringiensis serovar. kumamotoensis szerotípusra visszavezethető cry1A.105 (Cry1Ab, Cry1F és Cry1Ac „szintetikus” toxinkiméra). Idesorolható a B. thuringiensis serovar. tenebrionis szerotípusra visszavezethető m cry3A-gén, ami a cry3A módosított változata.
A Cry-toxinokat emlősökre, közöttük az emberre nézve többnyire ártalmatlannak gondoljuk. Immunszupresszált emberekben azonban gyulladásos folyamatokat indíthatnak el. Bacillus cereus által okozott bélhurut-járványokban észlelték a B. thuringiensis előfordulását is, aminek kivizsgálását elkezdték. A baktérium spórafehérjéinek mérsékelt allergizáló (nyálkahártya, bőr) hatását is leírták. Gyermekkori asztmával nem tudták összefüggésbe hozni. Ezek a hatások a legtöbb fehérjetípussal kapcsolatba hozhatók.
A Cry-toxinok közül a B. thuringiensis serovar. tolworthi cry9Ca1-gén kurtított változata által termelt Cry9c-toxint vádolták meg allergizáló hatással. Ennek az Aventis által 1997-ben engedélyeztetett fajtacsoportnak StarLinkTM (CBH-351) a neve. Több éves procedúra után visszavonták az élelmiszer célra való felhasználhatóságát és 2012 óta a dossziéjának karbantartása is szünetel. A Cry9c-toxint tartalmazó taco shell gyártása/felhasználása körül folyt a legtöbb vita. Tudományos szempontból ez is a lezáratlan viták egyike, ami a GMO-ügyek körül gyakori. Vagyis a bizonyítás egyik oldalról sem lezárt.
A negatív hatás bizonyítása a szabadalom tulajdonosa nélkül lehetetlen, mert egyszerűen nem adnak vizsgálati mintát, ha úgy gondolják, hogy a cégnek nem képezi érdekét a vizsgálat. Talán ezért mondják a géntechnológiát illetően elfogult, kevésbé tájékozottak azt, hogy a GM-növényekkel kapcsolatban eddig semmilyen tudományos aggályt nem sikerült bizonyítani. Valóban elképzelhető valakinek, hogy a nemzetközi cégek ok nélkül, bemondásra vonnak ki a piacról már engedélyeztetett genetikai eseményt, amely rendkívül komoly gazdasági befektetéssel járt?

26.ábra: A különböző Cry- és Cyt-toxinok hatásspektruma (Palma és mtsi nyomán)
Séralini és mtsai egy Monsanto tanulmány alapadatait újra elemezve a MON 863 (B. thuringiensis serovar. kumamotoensis eredetű preaktivált Cry3Bb1-toxint termel) esetében hepatorenális (májra és vesére kiterjedő tünetegyüttes) toxicitást vélt felfedezni. Az EFSA GMO Panelje nem értett egyet ezzel, de a közlemény körül széleskörű vita keletkezett. Tény, hogy jelenleg ennek a genetikai eseménynek az engedélye nincs meghosszabbítva Európában. Hasonló történt Kína és Fülöp-szigetek esetében, míg az Egyesült Államokban és Kanadában továbbra is engedéllyel rendelkezik a termesztése.
A nagy molekulatömegű Cry-toxinok kimutatására a környezetanalitikában ELISA-módszerek léteznek. Ezek használatával (Abraxis, Acadia, Agdia, Creativ Diagnostics, EnviroLogix) születtek nálunk Székács András (MTA Növényvédelmi Kutatóintézet, majd NAIK Agrár-környezettudományi Kutatóintézet) vezetésével mérési eredmények a MON 810 (Cry1Ab-toxin), a DAS-59122 (Cry34Ab1- és Cry35Ab1-toxin), valamint a Cry4-toxinokkal is. A legtöbb mérés a Cry1Ab-toxinnal végezték hazánkban (Takács Eszter és mtsai), amelyek azt mutatják, hogy a MON 810 kukoricalevél termeli a legtöbb toxint és ehhez képest keveset a szem. Ez jó hír a MON 810 szemet felhasználó baromfi- és sertéstartóknak, de vizsgálatokra kötelezné a csalamádé- és szilázs-felhasználók (szarvasmarha-, amur-tartók) körét.
Viszont, hogy a növény minden része termeli a Cry1Ab-toxint az azt jelenti, hogy nem szövetspecifikus, optimalizált kifejeződésről van szó. A MON 810 kukoricát kukoricamoly ellen fejlesztették ki, és részben bagolylepkék lárvái ellen. A kukoricamoly a szárban és csőben károsít, míg a gyapottok-bagolylepke csak a csövön. Míg a szárban kifejeződő toxinmennyiség maradéktalanul elpusztítja a lárvákat, addig a csőben kifejeződő kevesebb toxin erre csak hellyel-közzel képes. Ezért túlélők lesznek, amelyek Cry-toxinrezisztens rovarnépességet indítanak útjára (részletesen később). Miért termel Cry1Ab-toxint a gyökér, amellyel egyik hernyókártevő sem létesít kapcsolatot? Miért termelődik a Cry-toxin a pollenben, ami szétszóródik és szennyezi más lepkelárvák táplálékát (később részletesen)? Nem véletlen, hogy ezeket a korai fajtacsoportokat félkész-termékként nevezik meg azok, akik érdemben minősítenek.
A Cry1Ab-toxintermelés területegységre való vetítése eredményezett aztán igazán meglepő következtetést, mégpedig azt, hogy a MON 810 kukoricafajták a Dipel permetező szerben engedélyezetten kijuttatott mennyiség 50-1500-szoros mennyiségét termelheti meg egy hektáron (27. ábra). Ezen a ponton világossá válik, hogy ami engedélyezési szempontból egy terméknek tetszik (pl. MON 810), az a gyakorlat szintjén, konkrét fajtákra vonatkoztatva nagyon is különböző lehet környezetanalitikai szempontból.
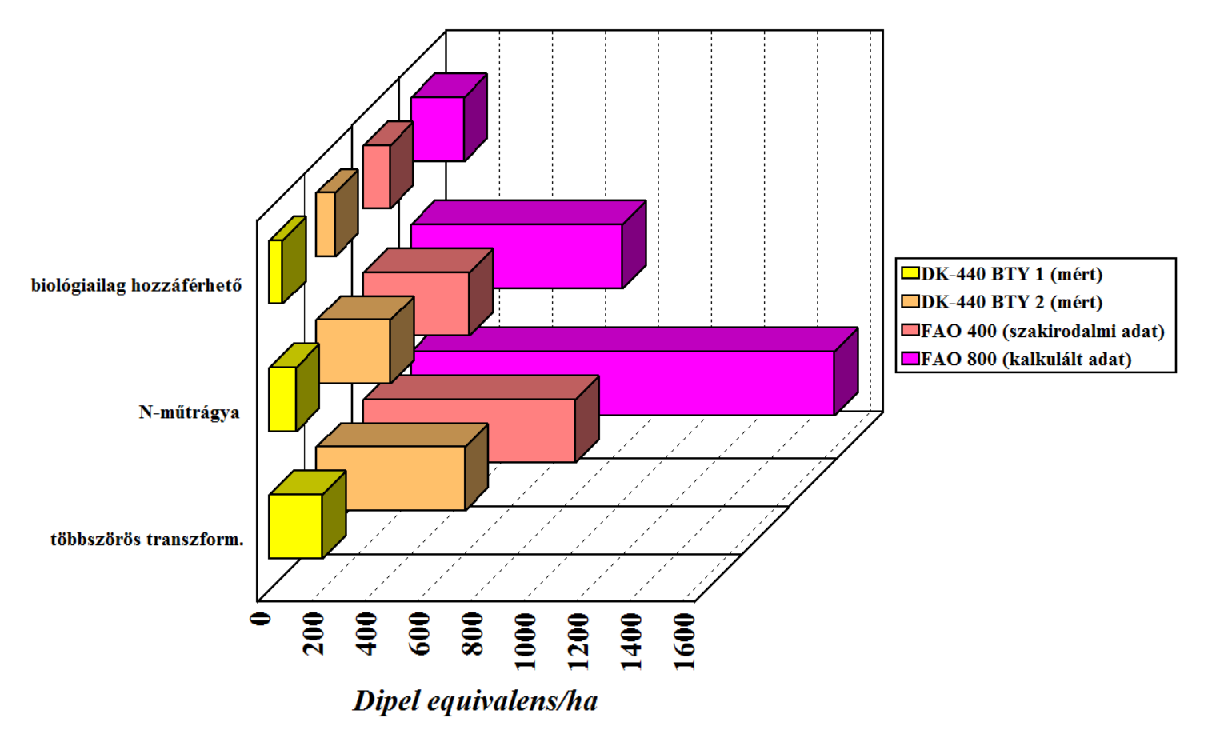
27.ábra: A DK-440 BTY (MON 810) Cry1-toxintermelése a Dipel-hez viszonyítva
A világon – így Európában is – az engedélyezés egy konkrét vegyület területi dózisához kötött. Az engedély megnevezi a kultúrát és azokat a készítménymennyiségeket, amire az engedély kiterjed. A munka- és élelmiszer-egészségügyi határértékek erre vannak kalkulálva, ettől eltérni nem lehet. A Cry-toxinok esetében más lenne a helyzet? Bizonyosan nem, a hatósági szabályozás nem tartalmaz ilyen jellegű megengedést. Az EPA/FDA és EFSA engedélyei jelenleg figyelmen kívül hagyják azt a tényt, hogy itt ennek a védekező anyagnak a sokszorosát a növény a területen termeli meg. Ilyen esetben az engedélyezést felül kellene vizsgálni, illetve ez csak úgy lehetne megengedő, hogy a Cry1Ab-toxinra vonatkozó területi dózist sokszorosára emelik. Ilyen azonban nem történt, s ez mindenképpen engedélyezési vakfolt.
A nagy mennyiségű Cry1Ab-toxin (tegyük hozzá MON 810 eredetű) megjelenése a termesztés területén tehát mérhető. Korántsem egyszerű viszont ez, mert az antitestet – az immunizálás során – a B. thuringiensis serovar. kurstaki protoxinjára nyerték (korrekció nélkül csak a bakteriális Cry1Ab- és Cry1Ac-protoxin mérésére alkalmazható), amely csak a keresztreaktivitás szintjén méri a kukorica preaktivált toxinját, vagyis a növényben aláméri (közel a felét mutatja csak), de a Dipel-ben sem pontos az eredmény, mert több toxinból (Cry1Aa, Cry1Ab, Cry1Ac, Cry2Aa) csak kettőt mér pontosan. Súlyos méréstechnikai kérdések merültek fel ebben az ügyben, amit az EFSA GMO Panelje a magyar tárgylóféllel folytatott megbeszélésén (53-67 old.) sem tagadott. Viszont a fajtatulajdonosnak nem kötelessége a termékellenőrzéshez szükséges megbízható mérőeszközök biztosítása? Miért nincs növényi Cry1Ab-preaktivált toxin mennyiségi vagy minőségi kimutatására alkalmas, optimalizált tesztcsomag forgalomban? Ugye, ebből is minden genetikai eseményhez eltérő kellene?
A folyamatosan termelődő és nagy mennyiségű Cry-toxin kizárja, hogy az integrált növényvédelem (környezetbarát védekezési eljárás, ami a kémiai védekezést sem zárja ki) területén alkalmazzák az ilyen GM-fajtákat, hiszen megelőző célú, nem okszerű védekezést biztosít (20-22 old.). A természetes B. thuringiensis készítményekkel szembeni védekezéshez képest itt két lényegi eltérést kell felsorolnunk. Az egyik, hogy nem a természetes Cry-protoxint termelik a növények, hanem annak egy géntechnológiai úton kurtított változatát (vö. preaktivált toxinfélék). Az ökológiai termesztés szigorúbb változatai már ez okból sem vennék be a Bt-növényeket a védekezési eszköztárukba, hiszen nem felel meg a felhasznált eszközökkel szemben támasztott „természetes” kritériumnak. Lehetne ezen hosszadalmasan vitatkozni, de felesleges. Ez a termesztési mód egyedi kritériumrendszert állított fel és a szerint termel, vásárlói ezt fizetik meg. A Bt-növény által folyamatosan termelt toxinmennyiség messze több (27. ábra), mint amit permetező szerként és okszerűen (ha szükség van rá) alkalmazunk. A mérgező hatás kialakulásában a dózisnak kulcsszerepe van, ahogy Paracelsus óta tudjuk.

81.kép: Kukoricatarló szártépés után (Fotó: Lauber Éva)
Az egy hektáron megtermelt nagy mennyiségű Cry1Ab-toxin biológiai hatása az, ami ezt követően érdekessé válik. A fajtatulajdonosok a Cry-toxinokkal kapcsolatos méréseket általában a B. thuringiensis bakteriális készítmények dossziéiból vették át. A Cry1Ab-toxin lebomlását például a talajba helyezett protoxinon mérték. Ezt nagyon gyorsnak találták. Más azonban a Bt-kukoricák tarlómaradványának sorsa, amit ma betakarítás után szártépővel kisebb részekre vágnak, majd a talajba forgatnak (81. kép).
Egy éven át nyomon követtük a tarlómaradványok sorsát és azt találtuk, hogy télen szinte egyáltalán nincs bomlás a növényi sejtekbe zárt Cry1Ab-toxintartalomban. Tavasszal kezdődik a bomlás, de a talajból mintázott növényi részekben egy év múlva is az eredeti Cry1Ab-toxinmennyiség 1-8%-át mérhetjük vissza. A 8% érték a fordulókban alakult ki, ahol nagyobb mennyiségű növényi törmelék került egy helyre. Ez tehát perzisztens maradékot jelent, ami a talajok tarlómaradvány-részeivel való felhalmozódást eredményezhet. Mindez két súlyos kérdést is felvet. Hogyan hat ez a talajban élő, aprító szerepű ízeltlábú közösségre, és miként befolyásolja ez az összetevő a talajban élő növénnyel szimbiotikus és növénymaradvány-bontó mikrobiális közösséget?
Kevés tudományos eredmény létezik arról, hogy a szerves anyag lebontását a talajban milyen tényezők befolyásolják, miközben kukoricában 5-6 tonna/ha tarlómaradványról beszélhetünk. Hazánkban Bakonyi Gábor végzett munkatársaival ilyen jellegű kísérleteket. Magyarországon a kukoricavetések talajában mintegy 200-300 állatfaj élhet. Ezek egymással és a mikroorganizmusokkal való együttműködése biztosítja betakarítás után a tarlón maradt szerves anyagok aprítását, ami a lebontás előfeltétele. Az eredményeik alapján a Cry-toxinokat tekintve nem érzékeny Folsomia candida ugróvillás a táplálkozása során különbséget tesz a Cry1Ab-toxint termelő és a közel izogénes vonal között. Ez egyfajta érzékszervi hatás, ami a Cry-toxinok érzékeny állatokon való tápcsatornai hatásától eltér. Két másik ugróvillás fajon (Heteromurus nitidus, Sinella coeca) viszont nem találtak hasonló következményt. Generációkon keresztül etetve ezt a módosított növényi törmeléket a F. candida népesség alkalmazkodása volt megfigyelhető. Ehhez tudni kell, hogy a száradó levélben a Cry1Ab-toxintartalom jelentősen lecsökken, és szoba-hőmérsékleten is mérhető lassú bomlás.
Az arbuszkuláris mikorrhiza kapcsolatban a gombák hifái áthatolnak a növényi gyökér kéregsejtjeinek sejtfalán. A gyökérben speciális képleteket (vezikulumok, arbuszkulumok) képeznek, melyeknek raktározó és felszínnövelő szerepük van. A szimbiotikus gomba-növény kapcsolat során a növény ásványi anyagokat (pl.: foszfort és nitrogént) és vizet kap a gombától, és cserébe kész szerves anyagokat juttat neki. A DAS-59122 (Cry34Ab1- és Cry35Ab1-toxinok) ennek a kapcsolatnak a kialakulását késlelteti a közel izogenikus vonalhoz képest. Hasonló mérési eredmény MON 810 (Cry1Ab-toxin) kukoricáról is ismert. Azt gondolom, hogy meghatározónak nevezhetjük még az ismerethiányunkat a talaj életközösségére gyakorolt hatást illetően.
A következő rész címe: A kiméra – Ikarusz röpte (Biotechnológikaland No32)
Darvas Béla
Bankszámlaszám: 12011265-01425189-00100001
Bank neve: Raiffeisen Bank
Számlatulajdonos: Átlátszónet Alapítvány
1084 Budapest, Déri Miksa utca 10.
IBAN (EUR): HU36120112650142518900400002
IBAN (USD): HU36120112650142518900500009
SWIFT: UBRTHUHB
Számlatulajdonos: Átlátszónet Alapítvány
1084 Budapest, Déri Miksa utca 10.
Bank neve és címe: Raiffeisen Bank
(H-1133 Budapest, Váci út 116-118.)
Támogasd a munkánkat az Átlátszónet Alapítványnak küldött PayPal-adománnyal! Köszönjük.
Támogatom PayPal-adománnyalHa az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
1% TÁMOGATÁS


Támogasd a munkánkat 10 ezer forint adománnyal, mi pedig megajándékozunk egy pólóval. Katt a részletekért.
Támogasd a munkánkat palackvisszaváltással, kattints az üvegvisszaváltós oldalra, mentsd el a kódunkat, és használd azt a Repontoknál!
Ha van bankkártyád, akkor pár kattintással gyorsan tudsz rendszeres vagy egyszeri támogatást beállítani nekünk az adjukossze.hu oldalán.


Postai befizetéssel is tudsz minket támogatni, amihez „sárga csekket” küldünk. Add meg a postacímedet, és már repül is a csekk.
Néző, Szurkoló, B-közép és VIP-páholy kategóriás Átlátszó-előfizetések között válogathatsz a Patreonon.
Bárhol is dolgozol a világban, ha a munkáltatód lehetőséget ad arra, hogy adott összeget felajánlj egy nonprofit szervezetnek, akkor ne feledd, a Benevity-n keresztül az Átlátszónet Alapítvány is ajánlható.
Ha az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
(Géntechnologikaland No78) Ukrajnában a GMO-k társadalmi megítélése összességében óvatos, sokszor elutasító. A közvélemény-kutatások és piaci tapasztalatok szerint a lakosság jelentős...

(Biotechnologikaland No77) A Marton Genetics Kft. állásfoglalása A Marton Genetics (Bázismag Kft.) cégcsoport a martonvásári kutatóintézet (Agrártudományi Kutatóközpont) munkájára épült....
(Biotechnologikaland No76) Előszó A GMO-lobbi hatására az Európai Bizottság javaslata (címábra) komoly visszhangra talált Európában. A javaslatok közül NGT1 esetében...

(Biotechnologikaland No75) Barabás Zoltán Biotechnológiai Egyesület kontra GMO-Kerekasztal A Barabás Zoltán Biotechnológiai Egyesület lépett hazánkban először abból a célból, hogy...
Támogasd a munkánkat banki átutalással. Az adományokat az Átlátszónet Alapítvány számlájára utalhatod. Az utalás közleményébe írd: „Adomány”, köszönjük!