Hétköznapi tudománypolitika. Alulnézet, ahol a csizma talpa a meghatározó élmény. Karcolatok a mezőgazdasági géntechnológiáról és az agrokemizálásról, tudományos hírek értelmezése
2010-től – miközben idehaza a kormányzati GMO-ellenes Road Show járta be a hazai felsőoktatást – a kutatásban megjelentek azok a géntechnológiai módszerek, amelyek a növényi géntechnológia egy haladottabb válfajának körvonalait kezdték kirajzolni. A mezőgazdasági tárca ekkorra meggyőzte magát arról – vagyis a tudományos tanácsoktól függetlenül önjáróvá vált –, hogy a GMO-ellenességgel szavazók tarthatók meg, így hát elégséges könnyed mosollyal azt mondani az udvariasan érdeklődő médiának, hogy „nem kell, mert nem kell”, megvédjük a magyar polgárok egészségét. Szinte kínos megjegyezni, hogy a magyar polgárok egészségét az Európai Unió élelmiszer-biztonsági rendszere (RASFF) szerint nem a GM-növények fenyegetik, hanem a táplálkozási eredetű patogének, a mikotoxinok és a növényvédő-szermaradékok. Ezen a szinten csak a glyphosate (vö. glyphosate-tűrő GM-növények) ügye köszön vissza ránk, de hazánk éppen ebben a megengedő tagállamok között sorakozott fel. Viszont mi a lényegük ezeknek az új géntechnológiai módszereknek, amelyekre egyre határozottabban mutogatnak? Valóban lényegileg más ez a korábbiaktól? Mostanában az MTA egyik mezőgazdasági bizottsága és egy pro-GM szemléletű civilszervezet GMO-párti Road Show-ja járja a felsőoktatási intézeteket. Mielőtt a genomszerkesztésbe alámerülünk, nézzük meg, hogy a kereskedelem által kínált termékek milyen megoldásokat kínálnak jelenleg, hogy ez alapján értékelhessük a haladást.
Minden történet sorsa az, hogy fordulóponthoz érkezik, és ami biztos, hogy nem az lesz a végső. A növényi géntechnológia számára is valami ilyesfélének jött el az ideje. A látszat ellenére nem annyira a technológia ebben a lényeg. A növényi géntechnológusok a kezdetektől tiltakoztak az ellen, hogy az előállítás technikája alapján ítéljék meg a fajtáikat. Azzal érveltek, hogy a terméket kell értékelni és valóban a környezet- és élelmiszerbiztonság szempontjából is ez a lényeges. Természetesen akkor, ha az előállításra használt technológia nem hagy maradandó nyomot (itt fajidegen örökítő anyagot) a termékben, ami akkor mégis elővigyázatosságra int. Próbáljuk ebből a nézőpontból végignézni az utat, mert a mostani fordulópont éppen arról szól, hogy elérkeztünk a genomszerkesztéshez (vagy legalább is ezt mondjuk róla) és annak mindjárt számos változatával is meg kell ismerkednünk.
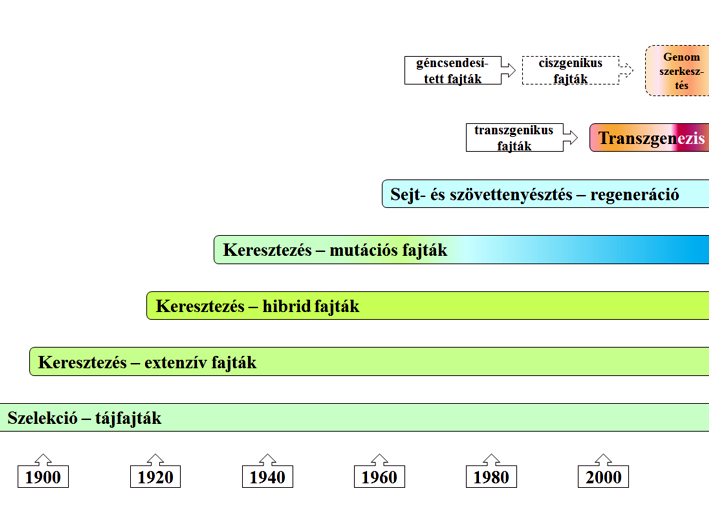
44.ábra: A nemesítés mérföldkövei (Jorasch, 2016 nyomán)
Miként is jelenik meg a növénynemesítés időben? Nos, kétségtelenül a szelekció (kiválogatás) volt az első nemesítési módszer, amit az ember alkalmazott, amikor rájött, hogy nem mindegy az, hogy milyen magot vet el. Igyekezett tehát a legjobban kinéző növények magjait eltenni a következő évi vetésre. Erre időszámítás (Krisztus születése) előtt tízezerből (vö. tönkebúza – Triticum dicoccum) vannak a legkorábbi adataink. Vagyis a tapasztalati alapú kiválogatás gyakorlata (pl. árpa, len, kukorica, köles, burgonya, repce, rozs stb.) messze megelőzte a genetika mendeli törvényeinek (1866) születését. Még az egyik legkésőbbi kultúrnövényünk, a cukorrépa szelekciója is hamarabb (1750) kezdődött. A kiválogatás szempontjai persze sokfélék lehettek, de valószínű, hogy a kiválasztott növény tulajdonságaiból megbecsült termésmennyiség lehetett a vezető elv. A rendszeres szelekció egyidejűleg a sokféleség ellen hat (118. kép), hiszen a bőtermő és ízletes fajták mellett más aligha foglalkoztathatta a korai nemesítőket. A nemesítési technikákkal bekövetkező génerózió és a vetőmag-világpiac kialakulásával rendkívüli mértékű tájfajtavesztés járt együtt. A ma hetvenfelé közelítő generáció bizonyosan emlékezik még arra az elképesztő alma és körte fajtaváltozatosságra, ami mára a múlté. Amit a század nemesítése nem karolt fel, az töredékeiben már csak a génbankokban létezhet.
A keresztezés lehetőségének felismerése már a genetika alapismereteinek birtokában következett be, majd követte ezt a hibridhatás felfedezése (44. ábra). A fizikai (röntgen-, gamma- és UV-sugárzás) és kémiai (pl. MMS, EMS, ENU stb.) eszközökkel indukált mutáció lehetősége (1930) követte mindezt, vagyis a fajta-előállítás eszközei és gyakorlata meglehetősen felgyorsult. A sikeres növényi sejt- és szövettenyésztés teremtette meg a lehetőségét az egy sejtből kiinduló növényregeneráció irányában (1960), bár kezdetben ezt a módszert elsősorban vírusmentes törzsanyagok (pl. burgonya, paprika stb.) előállítására használták. Gazdasági értelemben kétezer (az ISAAA statisztikája szerint 1996-ban) körül indult a szelekciós markerekkel segített transzgenikus fajtacsoportok előállítása és 2010-től itt állunk a genomszerkesztés új módszerei előtt, amiről beszélni fogok (lásd a következő részekben).
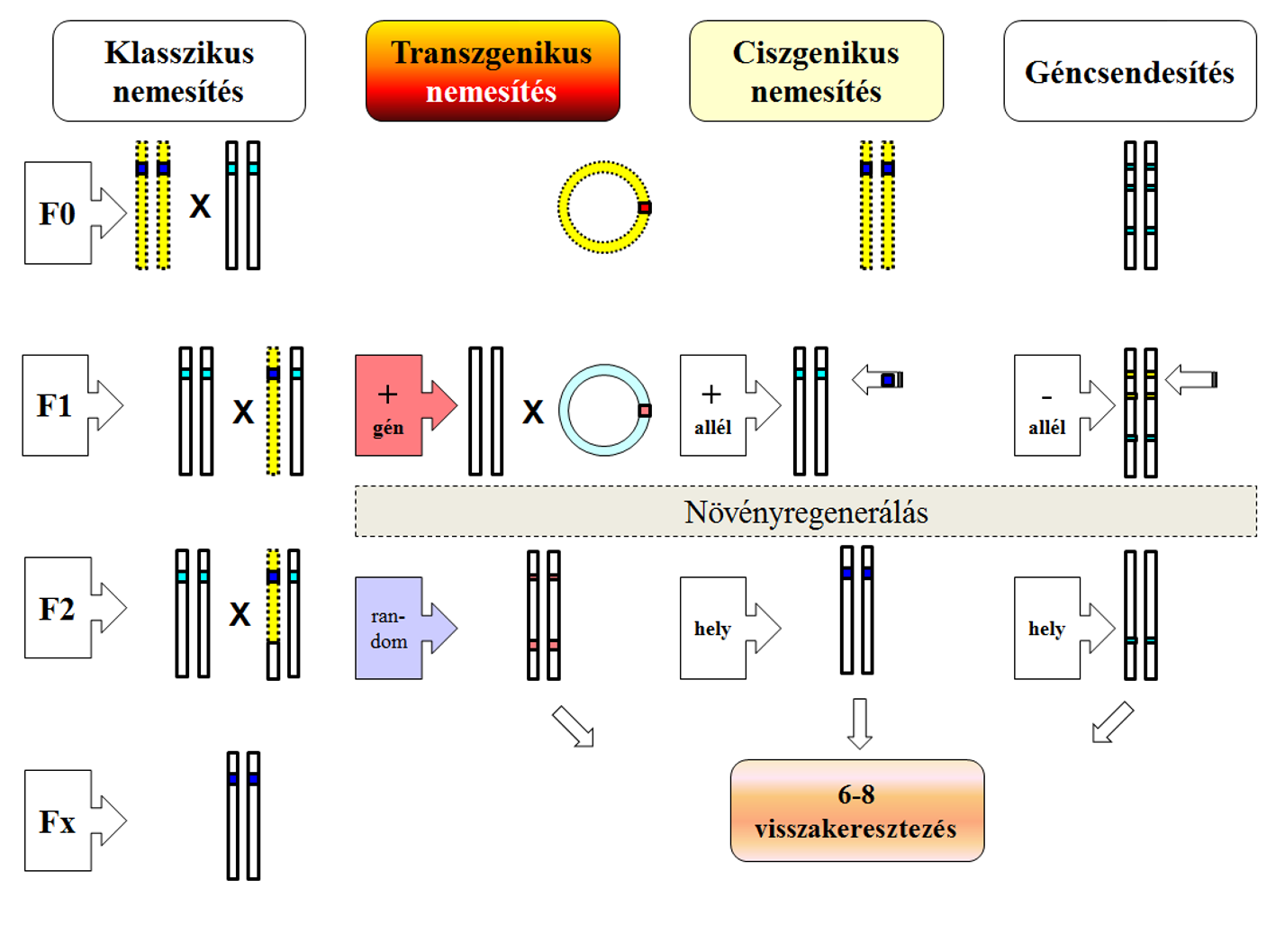
A transzgenikus és ciszgenikus növények elkülönítése az a szint, ami elérte a nem szakembereket is. Transzgenikus növények azok, ahol a faj genomjába egy másik fajból kerül be gén. Ez lehet növényi is, de ilyenkor spontán létrejöttére nincs esély. Az ezen az úton kapcsolatba kerülő fajok közötti távolság azonban ennél sokkal nagyobb lehet, hiszen bakteriális gének kerülhetnek át növényekbe. Közülük a Bt-növények a legismertebbek, amelyekbe a Bacillus thuringiensis nevű baktérium cry, vagy újabban vip génjei kerülnek be. Ilyen növények tehát a természetben nem jöhetnek létre, mivel ezeknek a géneknek (néha egyébként vektorszervezetek nagyon különös géncserét képesek végrehajtani) semmilyen útja nem ismert a növényi genom felé.
A ciszgenikus kifejezés eredetileg csak azt jelentette, hogy a gén (pl. mepsps, 2mepsps a kukoricában) előfordul az illető faj speciális népességeiben (faj alatt egységek: alfaj, változat, forma), vagyis a lehetséges allélek (természetes génváltozat) közül az egyik. Sőt közel rokon fajok is elfogadhatók voltak. Korábban ezt is elégségesnek tartották a fajtacsoport leválasztáshoz, ma azonban szűkebb értelemben akkor nevezünk egy fajtacsoportot ciszgenikusnak (ma még nincs ilyen a kereskedelemben, de az új genomszerkesztési technológiák ígérnek ilyet), ha az allél helyspecikus megjelenésű is, vagyis a kromoszómának azon a helyén/helyein fordulnak elő, amelyeken az illető növényben szokott (45. ábra). Ezen túlmenően a természetesnek megfelelő a promótere, az intronja és a terminátor génszakasza, amelyek sorrendje is azonos, vagyis a kifejezés a teljes működő génkontrukcióra vonatkozik úgy, hogy a géntechnológiai eljárásból (a transzformálásra használt plazmidból) semmilyen örökítőanyag nem marad vissza.
45.ábra: A molekuláris nemesítés vázlata
A gén eredeti helyét azért tartják többen fontosnak, mert ha a genomban nincs egy génnek helye, akkor elronthat egyet, aminek a helyébe kerül, illetve a környezetébe nem illő működése (a transzpromóterek általában bekapcsolt állapotúak) befolyásolhatják a normális életvitelt. Mások szerint az örökítő anyag másolása annyiféle hibával és javítgatással jár, amíg az egysejtű zigótából soksejtű egyedi élőlény válik, hogy az említett hatás akár el is hanyagolható és tulajdonképpen a mutációs hibák az okai az evolúciónak. Az európai szabályozás elővigyázatossági elve azonban nem ezt a fajta megengedő szemléletet támogatja.
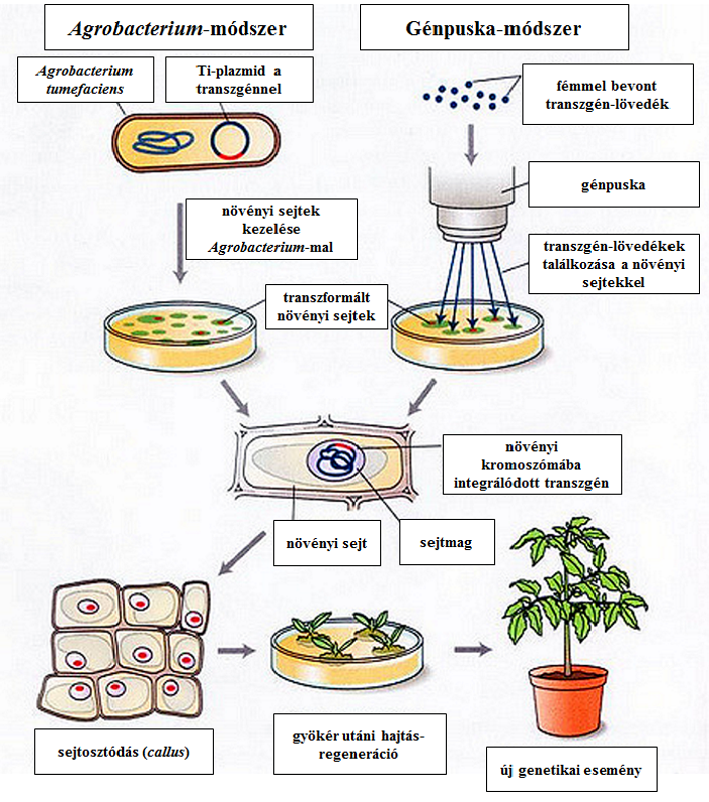
Az első generációs növényeknél alkalmazott véletlenszerű génbejuttatási módszerekkel (46. ábra) nem lehet (a valószínűsége nullához közelít) ciszgenikus fajtacsoportokat előállítani, hiszen a helyazonos allélcsere végrehajtására a génpuska, vagy az Agrobacterium-típusú vektorrendszer nem alkalmas. A génpuska különösképpen kritizált azért, hogy igen változatos helyre kerülhet a transzgén, s mellette transzgén-törmelékek is bejuthatnak a növényi genomba, mint ahogyan némely genetikai esemény eredeti dokumentációjában olvashatjuk. Ezekkel a beviteli módszerekkel létrehozott növények olyannyira egyediek és ismételhetetlenek, hogy megnevezésükre a genetikai esemény (genetic event) kódja (a fajtatulajdonos cég betűjele és számkombináció) szolgál. A genetikailag rendkívül pontatlan eljárásból (ezért sem helytálló a génsebészet kifejezés) létrejövő élőlényeket valójában a követő szelekció (ehhez a bevitt génkonstrukcióban valamilyen jól felismerhető markert alkalmaznak) segít a transzformáltak felismerésében és kiválogatásában. Ezt követően klasszikus szelekció (a fajta-előállítás időtartama tehát változatlan) választja ki azokat a növényeket, amelyek gazdasági célra használhatók. A lényeg, hogy egyidejűleg sok sejttel dolgozunk, ahol véletlenszerűen megvalósul az, ami lehetséges. Reménykedünk a véletlen jó találatban.
46.ábra: A növények transzformálásának vázlata
A fentiekből következik, hogy az eljárás során nem csupán a transzgén, hanem egy összetett vektorrendszer (több gén és génszakasz) kerül a növénybe, amelyben a részfeladatokat a különböző egységek oldják meg. Ez a transzformáltak felismerését is lehetővé teszi. A felismeréshez vagy DNS-vizsgálat (PCR), hiszen az idegen gének és részeik az illető növényben természetes körülmények között nem fordulnak elő, vagy a transzgén fehérjetermékének (pl. Cry-toxinok → ELISA) elemzése vezet el.
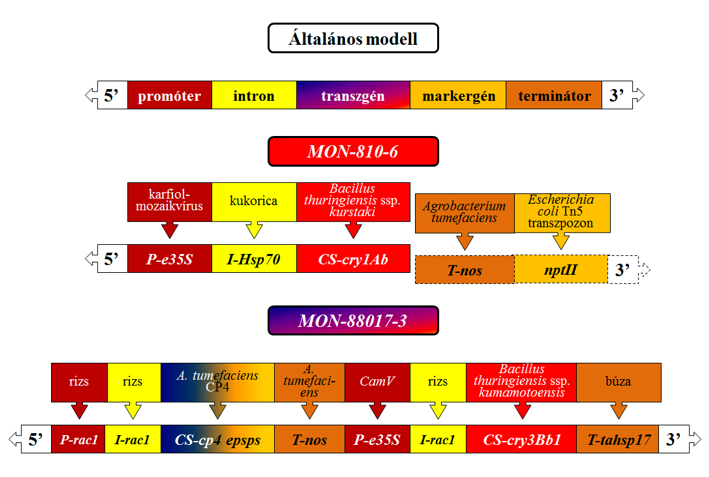
A génpuskával előállított fajtacsoportra a MON-810-6 (molyölő) példáján az alábbiak a jellemzőek. Itt kétféle plazmidot használtak a transzformációra: a PV-ZMBK07 és a PV-ZMGT10 nevűt. A PV-ZMGT10 plazmidon – az ISAAA adatbázisa szerint – markergénként glyphosate-tűrést előidéző géneket találunk (cp4 epsps, goxv247), viszont a CERA adatbázisa szerint ezek nem kerültek át a kereskedelmi forgalomba került fajtacsoportba (47. ábra). Az ISAAA adatbázisa jelzi, hogy a nptII szelekciós gént is tartalmaztak a transzformálásra használt plazmidok. A CERA adatbázisa leírja, hogy a kereskedelmi fajtákban CamV 35S karfiolmozaik-vírus eredetű promóter és Hsp70 kukorica eredetű intronfehérje génje mutatható ki. Azt is megtaláljuk, hogy a beépüléskor a 3’ vég leszakadt, vagyis a kereskedelmi fajtákban nem találunk több, a transzformáláskor felhasznált gént. Mindkét dokumentáció állítja, hogy a cry1Ab transzgén rövidített formája került a növénybe (45. ábra), ami így preaktivált Cry1Ab-fehérjetoxint termel. Azt mondhatjuk azonban, hogy a GM-fajták leírása szabadalmi okokból pontatlan és titkosított. A dokumentációból csak az kerül nyilvánosságra, amit a szabadalmi tulajdonos szeretne.
Az Agrobacterium vektorral (PV-ZMIR39 plazmid) végrehajtott módosítások közül egy több transzgénes változatot (MON-88017-3) választottam ki, ahol az egyik transzgén a szelekciós (glyphosate) marker szerepét is betölti (47. ábra). Az egyes transzgénekhez külön promóter, intron és terminátor tartozik. A cry3Bb1-gén ugyan Bacillus thuringiensis ssp. kumamotoensis-ből vált ismertté, de a transzformálásra használtat szintetikusan állították elő. A szintézis apróbb módosításokat tesz lehetővé.
A MON-810-6 példája is jól mutatja, hogy a plazmidot tartalmazó partikulum belövése, illetve az ezt követő kezelése a transzformáló genetikai tartalom sérülésével járhat, ugyanis a fajtacsoport-kifejlesztésére választott módosított egyedből már hiányzik (leszakadt) a terminátor és a neomycin/kanamycin-rezisztenciát biztosító rész (46. ábra). Hasonló eset nem ritka, hiszen a DAS-1507-1 fajtacsoport – amelyet úgyszintén génpuskával állította elő – tartalmaz egy ép és egy sérült cry1Fa2-gént is (45. ábra). Ez utóbbi promótere ugyan hiányzik, de stabilan öröklődött négy generáción át.
47.ábra: Két transzgenikus növényi génkazetta-vázlata (MON-810-6 – génpuska; MON-88017-3Agrobacterium-vektorral előállított több transzgénes módosítás, ahol cp4-epsps a szelekciós marker szerepét is ellátja)
A számunkra kívánatos tulajdonságai alapján kiválasztott transzformáns genetikai eseménynevet kap, amit szabadalmaztatnak (pl. MON-810-6, illetve kap védjegy-nevet is – ez esetben pl. YieldGard, MaizeGard), amivel forgalmazzák a fajtacsoportot. A genetikai esemény (pl. kukoricánál apavonal) nem azonos azzal, amit majd a vetőmag-cégnél a szerződés aláírása után átvehetünk. Ezt a GM-szülővonalat nagy teljesítőképességű fajtákkal (pl. DK-440) 6-8-szor visszakeresztezik, és így áll elő a módosított szülővonalra alapított fajtacsoport, ami a transzformáns tulajdonságait garantáltan örökíti az F1 generációban (pl. DK-440 BTY), miközben termőképessége és a helyi klímához való alkalmazkodása már a másik szülőre jellemző (45. ábra). Ez alapján érthető, hogy az egyes fajtacsoportokhoz tartozó konkrét fajták tulajdonságaiban nagyfokú eltérések lehetségesek, ami az azonos minőséghez szokott kutatókat igen sokszor megzavarja, amikor adataikat egymáséihoz viszonyítják. A későbbi nemzedékekben bekövetkező genetikai hasadás is az egyik oka annak, hogy a vásárolt GM-magból vetőmag nem fogható, vagyis azt minden évben meg kell vásárolni a fajtatulajdonostól, s megfizetni érte a technológia díjat. Ez már a hibridfajtáknál is így volt. A vetőmagot, mint terméket szabadalmi jog védi. A GM-technológia prominens cégei érték el először azt, hogy élőszervezetet szabadalmaztatni lehet, s ez egyúttal a nemzeti fajták védelme miatt Európában nagyfokú ellenállást váltott ki, ami máig tart. Ezen az úton ugyanis a nemzetek a szaporítóanyagra vonatkozó önrendelkezési jogukat elveszíthetik.
Miközben az elsőgenerációs GM-növényeket nagyon sokféle kritika éri és Európa országainak többsége (kivétel Spanyolország) nem mozdult arról az álláspontjáról, hogy területén nem tartja kívánatosnak a GM-fajtákat, a géntechnológiai módosítás módszertana nagy léptekkel haladt előre és szabadalmaztatott a korábbiaktól sokkal precízebb előállítási utakat. Ennek kapcsán korábban csak teoretikus kérdés volt az, hogy van-e lényegi különbség a genetikailag módosított és a géntechnológiai úton módosított szervezetek között. Különösképpen fontos ez hazánk számára, amely nem az Európai Unió nevezéktanát használta többségében a géntörvényben és a büntető törvényben. A géntörvény születésekor a Mezőgazdasági Biotechnológiai Kutatóközpontban szerveztek tanfolyamot, ahol „géntechnológiával módosított” elnevezés nyert.
Az alaptörvény fogalomhasználata az EU terminológiájával azonos: vö. „Az (1) bekezdés szerinti jog érvényesülését Magyarország genetikailag módosított élőlényektől mentes mezőgazdasággal…”. A géntörvény azonban az alábbi megnevezéseket használja „géntechnológiával módosított szervezetek”, „géntechnológiával módosított növények” stb, vagyis a kifejezésnek az eredeti angol nyelvűtől eltérő formáját alkalmazza. Előfordul azonban a géntörvényben is a „genetikailag módosított fajta” terminus (lásd 31§).
A Büntető Törvénykönyv is a „géntechnológiával módosított növényfajta” kifejezést használja. A hazai rendeletek, országgyűlési határozatok és kormányhatározatok többsége a géntechnológiai szót használja, csak egy országgyűlési határozat beszél „genetikailag módosított szervezetek”-ről, ami nemzetközi ügyet érint (Alpok-Adria kezdeményezés), így találkozhatott az EU tagországok eltérő szóhasználatával.
A terminológiai vita, amely a növényi genetikusok szerint tartalmi is a törvény előkészítő/karbantartó jogászokat kell, hogy hamarosan foglalkoztassa. Kérdés, hogy kettéválasztja-e a törvény ezt két fogalmat, aszerint, ahogyan ma használjuk (45. ábra). A géntechnológiával módosított vagy géntechnológiai úton módosított ez esetben is a genetikailag módosított (ennek természetes formái klasszikus növénynemesítési eljáráskor/keresztezéskor is hasonlatosak) jogilag megkülönböztetett alesetét képezné csak, de amit a törvény külön szabályozhatna. Ez utóbbi esetben szerintem a szabályozásnak a szűkebb értelemben vett ciszgenezishez (azonos helyen való allélcsere, amely a működést biztosító részekre is vonatkozik) való fogalomkötése nagyon is indokolt lenne, amelyet a mai növényi géntechnológusok a mutációs nemesítés folytatásaként aposztrofálnak.
A következő rész címe: Genomszerkesztés 2 – Helyspecifikus változatok (Biotechnológikaland No41)
Darvas Béla
Nélküled nincsenek sztorik.
Bankkártya
Átutalás
PayPal
1%
Így is támogathatsz
Támogasd a munkánkat banki átutalással. Az adományokat az Átlátszónet Alapítvány számlájára utalhatod. Az utalás közleményébe írd: „Adomány”, köszönjük!
Belföld
Külföld
Bankszámlaszám: 12011265-01425189-00100001 Bank neve: Raiffeisen Bank
Számlatulajdonos: Átlátszónet Alapítvány
1084 Budapest, Déri Miksa utca 10.
Ha az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
Támogasd a munkánkat 10 ezer forint adománnyal, mi pedig megajándékozunk egy pólóval. Katt a részletekért.
Üvegvisszaváltással
Támogasd a munkánkat palackvisszaváltással, kattints az üvegvisszaváltós oldalra, mentsd el a kódunkat, és használd azt a Repontoknál!
Bankkártyával az AdjukÖssze.hu oldalon
Ha van bankkártyád, akkor pár kattintással gyorsan tudsz rendszeres vagy egyszeri támogatást beállítani nekünk az adjukossze.hu oldalán.
Postai befizetéssel
Postai befizetéssel is tudsz minket támogatni, amihez „sárga csekket” küldünk. Add meg a postacímedet, és már repül is a csekk.
Havi előfizetés a Patreonon
Néző, Szurkoló, B-közép és VIP-páholy kategóriás Átlátszó-előfizetések között válogathatsz a Patreonon.
Benevity rendszerén keresztül
Bárhol is dolgozol a világban, ha a munkáltatód lehetőséget ad arra, hogy adott összeget felajánlj egy nonprofit szervezetnek, akkor ne feledd, a Benevity-n keresztül az Átlátszónet Alapítvány is ajánlható.
SZJA 1% felajánlásával
Ha az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
(Géntechnologikaland No78) Ukrajnában a GMO-k társadalmi megítélése összességében óvatos, sokszor elutasító. A közvélemény-kutatások és piaci tapasztalatok szerint a lakosság jelentős...
(Biotechnologikaland No76) Előszó A GMO-lobbi hatására az Európai Bizottság javaslata (címábra) komoly visszhangra talált Európában. A javaslatok közül NGT1 esetében...
(Biotechnologikaland No75) Barabás Zoltán Biotechnológiai Egyesület kontra GMO-Kerekasztal A Barabás Zoltán Biotechnológiai Egyesület lépett hazánkban először abból a célból, hogy...










Támogasd a munkánkat banki átutalással. Az adományokat az Átlátszónet Alapítvány számlájára utalhatod. Az utalás közleményébe írd: „Adomány”, köszönjük!