
Biotechnológikaland
Ukrajna és a GMO – Dakotarege
(Géntechnologikaland No78) Ukrajnában a GMO-k társadalmi megítélése összességében óvatos, sokszor elutasító. A közvélemény-kutatások és piaci tapasztalatok szerint a lakosság jelentős...


(Biotechnológikaland No38)
A Bt-növények általában 1-2 toxinfélét termelnek. A kártevők erre meglehetős gyorsasággal szelektálódnak. Kukoricabogár (Diabrotica sp.) lárvanépességeiben Cry3-toxin ellen, a kukoricamoly (Ostrinia nubilalis) lárváiban Cry1-toxin ellen egyaránt 6-7 éven belül alakul ki szabadföldi rezisztencia. Ez meglehetősen rövid idő a fajtacsoport-termékpályát tekintve. A Cry-toxinreceptorok keresztrezisztenciája alapján igyekeznek, ilyen kapcsolatban nem álló, eltérő toxint termelő variánsokat párosítani. A rezisztenciakezelés szabadföldi megoldásának ma még számos gyengesége van.

A Bacillus thuringiensis (Bt) cry-génjei által termelt Cry-toxinokról kiderült, hogy rendkívül sokfélék. Nagyságuk és ún. konzervatív szakaszaik helyzete is eltérő lehet, miközben hatásspektrumuk is változhat (39. ábra). Még csak nem is az egyetlen toxinféléről van szó (vö. bin, cyt, mtx, sip, vip), amelyet ez a talajlakó/rovarpatogén baktériumfaj termel (40. ábra), s amelynek plazmidon lévő cry-génjeit esetleg rokonának, a Bacillus cereus patogén törzseinek konjugáció során kölcsönadhatja. Korántsem minden szempontból gyanú fölött álló baktériumról és termékeiről van szó. Hanem olyanról, ami úgy tűnik, hogy változatosságában számtalan meglepetést tartogat a számunkra. Mivel a Bt-növények ezeknek a géneknek a kurtított változatait tartalmazzák (ezek az ún. preaktivált toxinokat termelik), ezért az alapos környezet-egészségügyi ellenőrzésük nagyon is indokolt, vagyis nem lehet olyan egyszerűsített módon tekintetni rájuk, mintha aggályoskodásra alapot nem adna ez a fehérjetoxin-csoport.
Három, a Bt-törzsekkel is kapcsolatba hozott, mikroorganizmusok által termelt, közel rokon toxincsoportot ismerünk (40. ábra). Ezek egyike a binary toxinok csoportja (szélesebb körben pórust formáló toxinokként ismertek), amelyek közül a lépfene (Bacillus anthracis) toxinja a legismertebb (bár termelik a Clostridium botulinum, C. difficile, C. perfringens és C. spiroforme törzsei is), s amely a biológiai hadviselés figyelmét is felkeltette. A parasporin-okhoz tartozó toxinokat a B. cereus csoport tagjai (B. cereus, B. anthracis, B. mycoides, B. pseudomycoides és B. weihenstephanensi) – termelik. A ricinusból leírt ricin (lektin) viszont a Bin- és Cyt-toxinokkal mutat részbeni hasonlóságot.
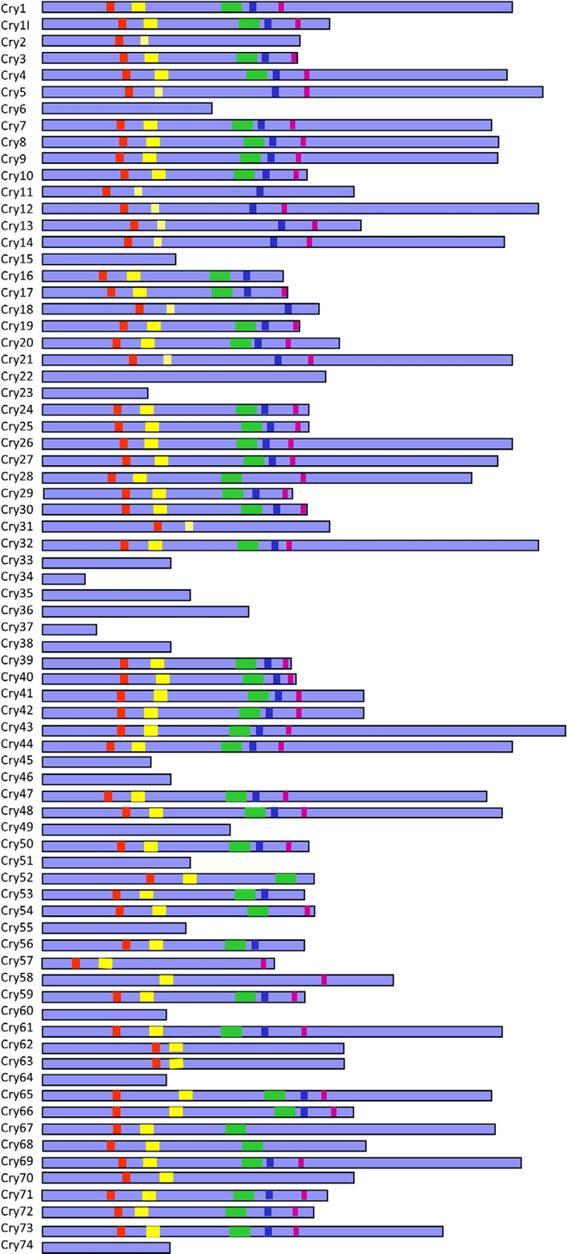
39.ábra: Cry-toxinok nagysága és öt konzervatív régiójának helyzete (Forrás: Adang és mtsai)
A Bt-növények kurtított cry-géneket tartalmaznak. Ezek a gyakorlatban maradt kurtított gének vagy a cry1 (cry1Ab, cry1Ac, cry1Fa2), cry2 (cry2Ab2), cry3 (cry3A, cry3Bb1, cry34Ab1, cry35Ab1), cry9 (cry9C) csoportba, vagy az ugyanezen baktérium által termelt vip [vip3A(a), vip3Aa20] gének közé tartoznak. Léteznek ma már szintetikus cry génváltozatok is, pl. a cry1A.105 (Cry1Ab-, Cry1F- és Cry1Ac-toxinhatás), mcry3A és az ecry3.1Ab (Cry3A- és Cry1Ab-toxinhatás).
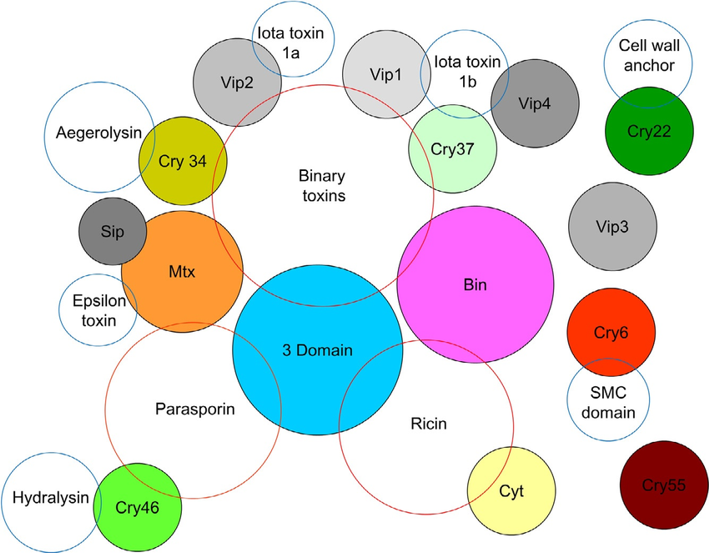
40.ábra: Cry-toxinok és rokonsági körük (színezett körök) – a kék kontúr a nem Bt-eredetű -toxinokkal való szerkezeti rokonságot jelöli, míg a piros kontúr közös részekre (alegység) utal a binary-, parasporin– és ricin-típusú toxinokkal (Forrás: Adang és mtsai)
A Cry-toxinok rendszintű specifitást mutatnak a rovarok között, vagyis a Cry1-toxinok a Lepidoptera hernyók, míg a Cry3-toxinok a Coleptera lárvák ellen hatnak. Nincs hatásuk az imágókon, s ez a tulajdonságukat védekezéstechnikai szempontból fontos megjegyezni. A Cry2-toxint a Cry1-toxinra rezisztenssé vált hernyónépességek ellen szokták bevetni. A Cry9-toxin szintén Lepidoptera-specifikus, de allergén gyanú miatt az iparilag fejlett országok kínálatában már nem szerepelnek. A gyakorlatba került Vip-toxinok szintén Lepidopera-specifikusak.
A nyilvántartó adatbázisok közül az International Service for the Acquisition of Agri-biotech Applications (ISAAA) adatbázisa a legrégebbi, és adatfeldolgozását mindig is az ipar igényei szerint végezte. Erre a pro-GM szemléletű adatfeldolgozásra jellemző a trait hektár fogalma, amit úgy számítanak, hogy egy többszörös módosítást tartalmazó genetikai esemény (stacked events) többszörös területfoglalással tüntetik fel, vagyis az általuk megadott termesztési adatok felfelé torzítanak. Más tekintetben az adatbázis korrekt, bár európai viszonylatokra nézve mindig is ellenmondásos volt. A támogatás hiányában bezárt GMO-Compass (amely az európai fogyasztóknak készült nyilvános adatbázis volt) az Európai Unió viszonylatait másként mutatta. A pillanatnyilag elérhető EU regisztrációs rendszer igen kevés adatot tartalmaz, a korábbiakhoz képest. Lényeges, hogy a világ országaiban eltérő nemzeti stratégiával viszonyulnak a GM-növényekhez.
2015-re az egyszeresen módosított „gyomirtószer-tűrő” (pl. glyphosate-tűrő) eseményekhez tartozik a termesztési adatok 53%-a, a Bt-növényekhez a 14%, míg 33%-a többszörös módosítások (pl. glyphosate-tűrés + kukoricabogár elleni védelem) kategóriájába esik. Azt mondhatjuk, hogy a Bt-növények gazdasági sikere messze a glyphosate-tűrő növények mögött kullog. Sőt, ezen a területen belül is, 2015-re a többszörösen módosítottak (60 millió hektár) dominálnak az egyszeresen módosított (20 millió hektár) fajtacsoportok (genetikai események) fölött (41. ábra). Az észak-amerikai választékban a Bt-növények sokkal inkább a többszörös genetikai eseményekben, mint technológiát kiszélesítő tulajdonságok jelennek meg. Az európai engedélyezésben már két olyan eseményt találhatunk, amelyek nem szerepelnek önállóan az ISAAA adatbázisában. A BCS-GH4-7 gyapot és a MON-87701-2 szója önállóan nem elérhetőnek van jelölve, illetve csak többszörös eseményben partnerek.
Csak az egyszeres eseményeket (35 Bt-esemény) elemezve burgonyában 16, padlizsánban 1 esemény található az adatbázisban, míg egy sem az EU regisztrációs rendszerében. Gyapotban és kukoricában az ISAAA adatbázisában több esemény található, mint az EU-ban bejelentésre került. Ez azt mutatja, hogy az európai engedélyezés egyre jelentősebb fáziskésésben van. Mindez csak részben származhat az elővigyázatosság elvéből, hiszen az import, a feldolgozási, a takarmányozási és élelmiszer- felhasználási engedélyek területén az EU is meglehetősen engedékeny. Ez arra utal, hogy az Európai Élelmiszer-biztonsági Hatóság (EFSA) nem ad érdemi hitelt az elmarasztaló dietétikai kritikáknak. Nem így van ez a vetést illetően, ahol eddig csak a MON-810-6 genetikai esemény szerzett engedélyt, de az EU tagállamai közül többen is vetési moratóriumot léptettek életbe.
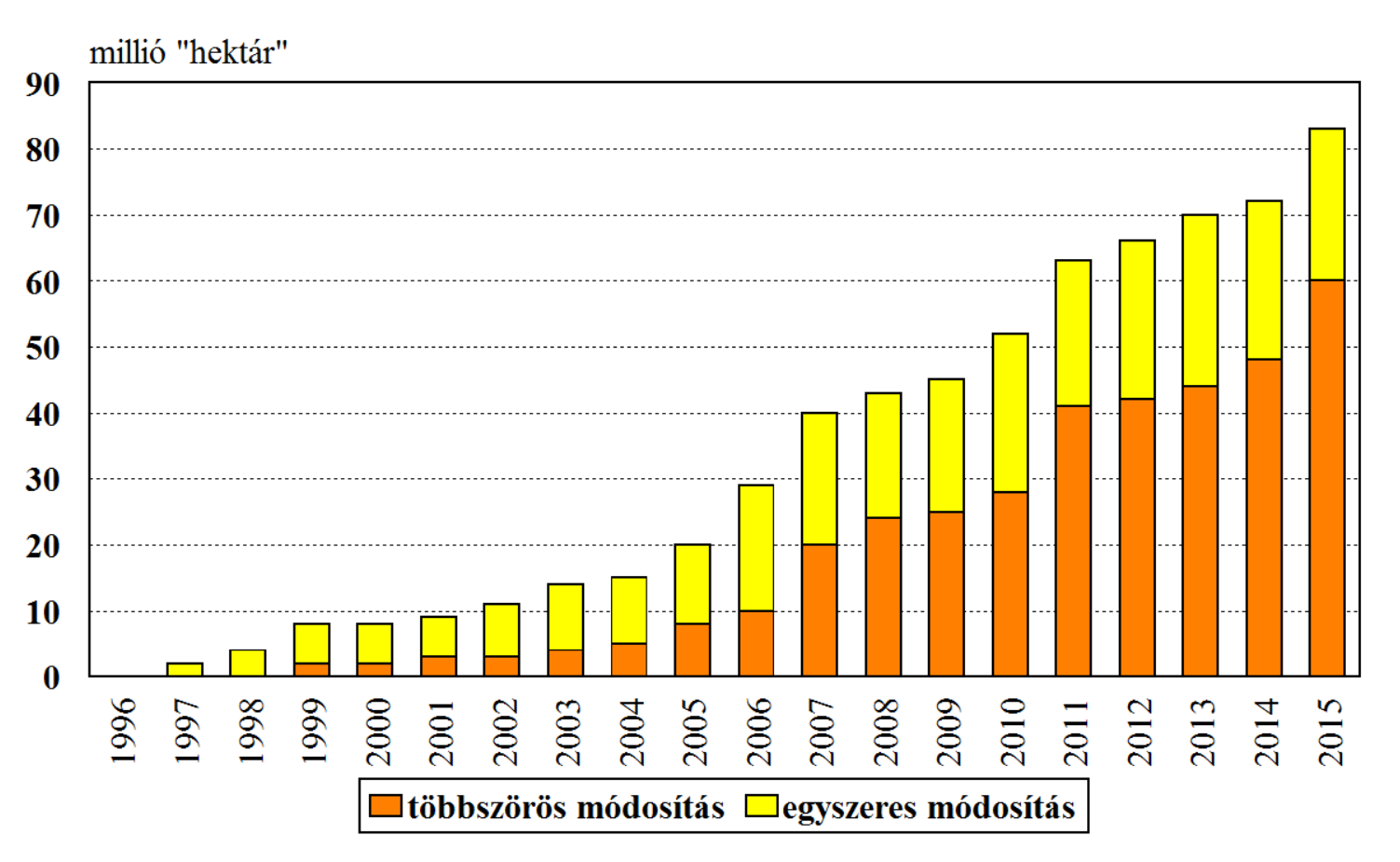
41.ábra: A Bt-növények vetésterülete 1996-2015 között (az ISAAA trait hektár adatai alapján)
A transzgenikus Bt-növényeket arra a célra fejlesztették ki, hogy rezisztenciát mutassanak a kultúrnövények kártevőire. Hamarosan megjelentek azonban az ilyen növényekre tűrőképességet mutató kártevő-népességek. A bulvársajtó ezeket hívja helytelenül „szuper-kártevőknek”. Az ismeretterjesztő sajtó ezt a jelenséget esetenként hatalmas tudományos újdonságként kezeli, azonban már az eddigiek szerint is bizonyosan állítható, hogy nem az tudományos szenzáció, hogy a rezisztencia megjelenik, hanem az ellenkezője lenne, hogy az érintett élővilág nem képes alkalmazkodni. Az evolúció során a biodiverzitás részeként nagyfokú válaszképesség alakult ki. Ez az oka annak, hogy ökológusok a biodiverzitást kulcsfontosságú szerepűnek látják az emberiség jövőjét tekintve is.
Cry1-toxintermelő, lepkehernyó-rezisztens növények
A tévesen „rovar-rezisztensnek” hívott Bt-növények a termelt Cry-toxintípus alapján valamely rovarrend ellen biztosítanak védelmet (vagyis valójában hernyó-rezisztensnek kellene nevezni a Cry1-toxintermelőket). Számunkra a kukorica-kártevői a legfontosabbak, amelyek közül a tűzmolyok családjába (Crambidae) tartozó európai kukoricamoly (Ostrinia nubilalis) a világon általános előfordulású, a csöveken károsító további hernyó-kártevők pedig már a bagolylepkék (Noctuidae) családjából kerülnek ki. A Helicoverpa zea (címkép) észak-amerikai, a Spodoptera frugiperda dél-amerikai, a Helicoverpa armigera kelet-európai (112. kép), míg a Sesamia nonagrioides nyugat-európai kártétele emelhető ki ezek közül. Mindez ugyanazon genetikai esemény főhatás-megítélését az eltérő kontinenseken különbözővé teszi.

112.kép: A gyapottok-bagyolylepke (Heliothis armigera) hernyója csicseriborsó hüvelyében (Fotó: J. Wessels)
Korábban úgy vélték, hogy a természetes eredetű vegyületekre kisebb eséllyel és lassabban alakul ki rezisztencia, mint a szintetikus vegyületekre. Ezt azonban a penicillin esete elég korán cáfolta. A tárgyalt téma szempontjából viszont idetartozó, hogy a rovarirtó hatóanyagokkal kapcsolatos rezisztenciairodalom vezető faja a Helicoverpa armigera, amelyben 2008-ig 435 dokumentált esetet tartanak nyilván.
A MON-810-6 esetében a cry1Ab-gén került be a kukorica genomjába. Lényegi különbség, hogy míg a baktérium mindig többféle toxint termel, és azt protoxin formájában tartalmazza, addig a Bt-növény csak 1-2 félét termel. Rezisztencia szempontjából tehát az eseteink az „egy- és többszűrős” eltérő csoportokba tartoznak. Vizsgálatok szerint a rezisztencia kialakulásnak esélye 300-szor nagyobb egy, mint négy Cry-toxinkeverék esetében.
2001-ben, a MON-810-6 eseményből származó DK-440 BTY Bt-kukoricalevél-őrleménnyel, aszalványmoly-lárvákon elvégzett kísérletben már az első három nemzedékben megfigyeltük a túlélők számának növekedését. A 10. generációban fele tömegű bábok fejlődtek, bár az embrionális + posztembrionális kifejlődési időtartamuk közeledett a kontrolltápon tartottakéhoz. A 20. generációban nem tapasztaltunk mortalitási különbséget és az izogenikus kukoricalevél-őrleménnyel kevert tápon fejlődő állatokhoz hasonló volt a bábtömeg és a kifejlődési idő is. A 30. nemzedékben vizsgáltuk, hogy mi történik a Cry1Ab-tűrő törzsünkkel, ha az Dipel-lel találkozik, amiben nagyobb mennyiségben ötféle protoxin van jelen. A rezisztens törzsünk közel négyszer volt Dipel-kezelésre kevésbé érzékeny. Ez keresztrezisztenciát jelez. Egy kísérleti variánsban tíz nemzedékre felfüggesztettük a Cry1Ab-toxint tartalmazó levélőrlemény adását, azonban a rezisztencia fennmaradt, ami szerint a tenyészetünkben kialakuló rezisztencia öröklődő természetű.
Bár laboratóriumokban már előzetesen is jutottak 160-520-szor tűrőképesebb népességekhez, Alves és mtsai 2006-ban publikálták eredményeiket két rezisztens Ostrinia nubilalis törzsről, amelyek Cry1Ab-toxinnal kapcsolatos érzékenységcsökkenése 1300-2000-szeres volt. A rezisztencia öröklődő természetű, és az ez esetben több gén megváltozott aktivitására volt visszavezethető. A rezisztenciához vezető fő gén előfordulását az észak-amerikai vad kukoricamoly-népességekben csekélynek gondolják, s ez az oka annak, hogy ellenállóság a gyakorlatban lassabban alakul ki a vártnál. Ettől eltérően a Cry1Ab-toxint termelő törzsek 1996-ban köztermesztésben való megjelenése után hat évvel, 2001-ben Crespo és mtsai Kandiyohi-ban (Minnesota) szabadföldről izoláltak rezisztens O. nubilalis-t, amely a későbbi szelekció után közel 800-szor volt ellenállóbb Cry1Ab-toxinra.
Bt-gyapotból (Egyesült Államok, 2005) Cry1Ac- és Cry2Ab-rezisztens Helicoverpa zea; Bt-kukoricából (Puerto Rico, 2006) Cry1 F-rezisztens (DAS-1507) Spodoptera frugiperda, és Cry1Ab-rezisztens (MON 810, SYN-Bt11) Busseola fusca népességeket (Dél-Afrika, 2006) írtak le. A Cry-toxinrezisztenciának többféle oka lehet: az emésztőenzimek (tripszin, kimotripszin) aktivitás- vagy összetétel-változása; hatékonyabb detoxifikáció (citokróm P-450 és glutation-S-transzferáz enzimkészlet-eltérések); a bélben lévő Cry-receptorok számának vagy érzékenységének eltérései; végül az eredményesebb sebregeneráció. Több Cry1Ab-rezisztens kukoricamoly-törzsnél azt találták, hogy a középbél membránjának felépítésében résztvevő cadherin szerkezete eltér a normálistól, amit egy mutáns cdh gén okoz.
Hazánkban kétszáz fölötti védett és fokozottan védett lepkét tartunk számon. A csalánféléken táplálkozó atalantalepke (Vanessa atalanta), c-betűs lepke (Polygonia c-album), kis rókalepke (Aglais urticae) és nappali pávaszem (Inachis io); szederféléken (Rubus sp.) ibolya gyöngyházlepke (Argynnis niobe), lápi gyöngyházlepke (Brenthis ino), nyugati törpebusalepke (Spialia sertorius) és zöldes gyöngyházlepke (Argynnis pandora) a leginkább érintettek. Az eddigi méréseink az I. io lárváin arra utalnak, hogy ezeknek a népességeikben is találhatók rezisztens egyedek.
Cry3-toxint termelő, bogárlárva-rezisztens növények
Az Egyesült Államokban (és onnan való betelepülése után már Európában is) a kukoricabogár-fajok (Diabrotica spp.) a kukorica legjelentősebb kártevőjévé váltak. Bogárlárvák ellen a Bacillus thuringiensis Cry3-toxinokat termelő patovariánsai (tenebrionis, kumamotoensis) alkalmasak (113. kép), tehát a növényi géntechnológia ilyen gének felé fordult. A kukoricabogarak lárvái a gyökeret károsítják, ebben a növényi részben azonban – az eddig ismert változatokban – a cry3-gén kifejeződése szerény.

113.kép: Kukoricabogár (Diabrotica sp.) lárvájának gyökér-kártétele. Balra Cry3Bb1-toxint termelő törzs, jobbra károsított közel izogenikus vonala (Fotó: C. Kruppke).
Fentiek következménye, hogy már az első éves alkalmazásnál sem teljes a hatás, tehát vannak túlélők. Ezt azt eredményezi, hogy a szubletális hatás miatt a szelekció erőteljes és a Cry3-rezisztencia kialakulásának rizikója itt különösképpen nagy. Ennek megfelelően nem meglepő, hogy a 2003-ban kezdődő termesztés utáni hetedik évben, 2009-ben Iowa körzetében Gassmann és mtsai szabadföldi körülmények között kialakult Cry3Bb1-toxin rezisztens Diabrotica virgifera virgifera népességekről számoltak be. A Cry3-rezisztens kukoricabogár-népességek ellen a cry34Ab1 + cry35Aa1-géneket tartalmazó kukorica sem mutatott számottevő ellenállást.
A Cry-toxinrezisztencia kezelése
A Bt-növényeken szelektálódó kártevők miatt a rezisztenciakezelés (menedzselés) a kezdetektől fogva fontos gyakorlati kérdés. Az egyik megközelítés abból indult ki, hogy a több Cry-toxint tartalmazó készítményekkel szemben az ellenálló képesség lassabban alakul ki. Napjainkban tehát több cry-gént tartalmazó fajták terjednek. A Cry-receptorok elkülönítése során vizsgálták azt, hogy azokhoz mely Cry-toxinok kötődnek, és ilyen módon a keresztrezisztenciára vonatkozó ismereteket is szereztek. A káposztamoly (Plutella xylostella) lárvákból izolált Cry-receptorok esetében a Cry1C-toxin keresztérzékenységet mutatott CrylAb-, CrylAc- és Cry1F-toxinokkal, de közepes volt a keresztérzékenység Cry1Aa- és Cry9C-toxinok esetében. Keresztérzékenységet nem találtak Cry1Bb-, CrylJa- és Cry2A-toxinok vizsgálatakor. Keresztérzékenység a II domén-szerkezetével volt kapcsolatba hozható (42. ábra).
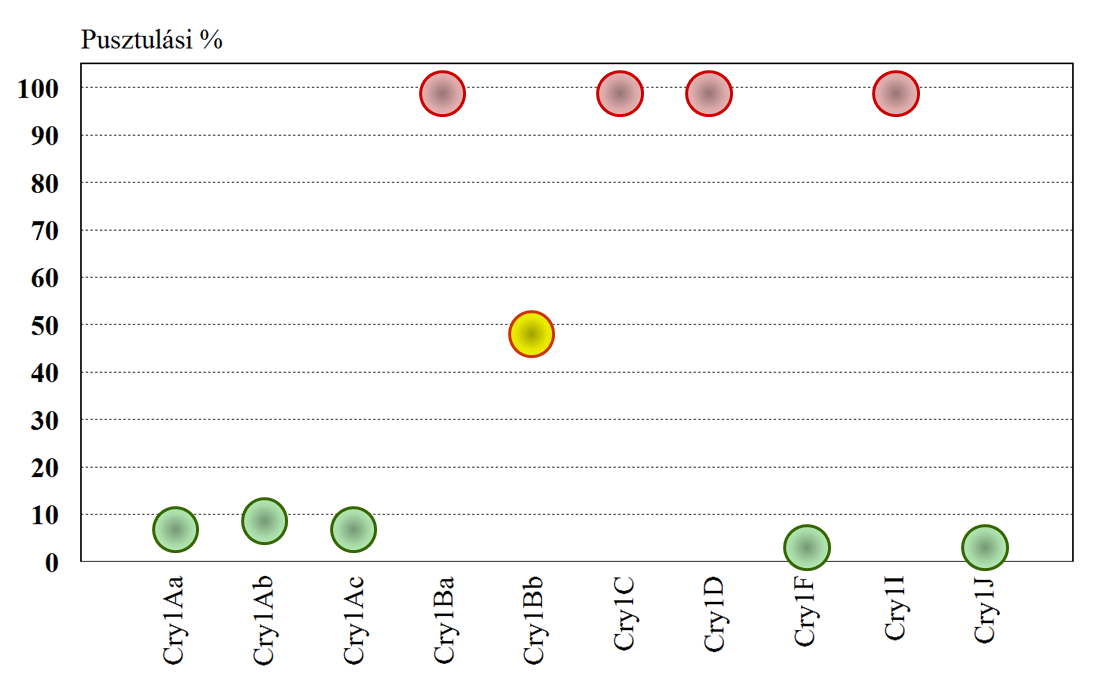
42.ábra: Cry1A-toxin-ra rezisztens káposztamoly lárvákon százszoros koncentrációban adott Cry1-toxinok hatása (Forrás: Tabashnik és mtsai)
A menedzselés másik útja a Cry-rezisztencia recesszív módon öröklődő természetére támaszkodik, vagyis arra a jelenségre, hogy ha egy Cry-toxinra érzékeny egyed egy rezisztens egyeddel párzik, úgy az utódok elég érzékenyek lesznek ahhoz, hogy a Cry-toxin tartalmú kukorica elpusztítsa azokat. Ezért javasolják azt, hogy a Cry1- és Cry3-toxinokat tartalmazó fajtacsoportok esetében kb. a terület 10-20%-át elérő területen Cry-toxint nem termelő kukoricából ún. „menedékzónát” létesítsenek. Ezt teremt arra esélyt, hogy a nem módosított táblákban kifejlődő érzékeny egyedek a túlélő Cry-rezisztens egyedekkel párosodjanak.
Ennek a gyakorlatnak azonban számtalan gyöngesége van: (i) a terület jelentős részén tenyészti a kártevőt; (ii) a menedékzóna egymással érintkező soraiban Cry-toxin tartalmat illetően vegyes szemeket tartalmazó csövek jönnek létre, amelyeken a kártevő túlélésének valószínűsége megnő; (iii) a Cry-toxint tartalmazó növényeken a kifejlődés sebessége jelentősen lelassul a menedékzónában fejlődő rovarokhoz képest, ugyanis a szubletális hatás egyik jelentős megnyilvánulása a lárvális fejlődési idő közel kétszeresre való megnyúlása. Hazánk agroklimatikus körülményei mellett ez azt jelenti, hogy az Ostrinia nubilalis és Helicoverpa armygera első nemzedékeinél az érzékeny és a rezisztens népességek egyedei jelentős mértékben nem is pározhatnak egymással, hiszen a rezisztens egyedek párzására az érzékenyekénél sokkal később kerül sor, vagyis a két népesség a párzásban elkerüli egymást. Csupán az áttelelő népességek esetében van esély arra, hogy az áttelelés utáni tavaszi bábból való kibúvás hasonló időben történjen.
A következő rész címe: III. Országgyűlési Nyílt Napok a GMO-król – Rendhagyó előszó (Biotechnológikaland No39)
Darvas Béla
Bankszámlaszám: 12011265-01425189-00100001
Bank neve: Raiffeisen Bank
Számlatulajdonos: Átlátszónet Alapítvány
1084 Budapest, Déri Miksa utca 10.
IBAN (EUR): HU36120112650142518900400002
IBAN (USD): HU36120112650142518900500009
SWIFT: UBRTHUHB
Számlatulajdonos: Átlátszónet Alapítvány
1084 Budapest, Déri Miksa utca 10.
Bank neve és címe: Raiffeisen Bank
(H-1133 Budapest, Váci út 116-118.)
Támogasd a munkánkat az Átlátszónet Alapítványnak küldött PayPal-adománnyal! Köszönjük.
Támogatom PayPal-adománnyalHa az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
1% TÁMOGATÁS


Támogasd a munkánkat 10 ezer forint adománnyal, mi pedig megajándékozunk egy pólóval. Katt a részletekért.
Támogasd a munkánkat palackvisszaváltással, kattints az üvegvisszaváltós oldalra, mentsd el a kódunkat, és használd azt a Repontoknál!
Ha van bankkártyád, akkor pár kattintással gyorsan tudsz rendszeres vagy egyszeri támogatást beállítani nekünk az adjukossze.hu oldalán.


Postai befizetéssel is tudsz minket támogatni, amihez „sárga csekket” küldünk. Add meg a postacímedet, és már repül is a csekk.
Néző, Szurkoló, B-közép és VIP-páholy kategóriás Átlátszó-előfizetések között válogathatsz a Patreonon.
Bárhol is dolgozol a világban, ha a munkáltatód lehetőséget ad arra, hogy adott összeget felajánlj egy nonprofit szervezetnek, akkor ne feledd, a Benevity-n keresztül az Átlátszónet Alapítvány is ajánlható.
Ha az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
(Géntechnologikaland No78) Ukrajnában a GMO-k társadalmi megítélése összességében óvatos, sokszor elutasító. A közvélemény-kutatások és piaci tapasztalatok szerint a lakosság jelentős...

(Biotechnologikaland No77) A Marton Genetics Kft. állásfoglalása A Marton Genetics (Bázismag Kft.) cégcsoport a martonvásári kutatóintézet (Agrártudományi Kutatóközpont) munkájára épült....
(Biotechnologikaland No76) Előszó A GMO-lobbi hatására az Európai Bizottság javaslata (címábra) komoly visszhangra talált Európában. A javaslatok közül NGT1 esetében...

(Biotechnologikaland No75) Barabás Zoltán Biotechnológiai Egyesület kontra GMO-Kerekasztal A Barabás Zoltán Biotechnológiai Egyesület lépett hazánkban először abból a célból, hogy...
Támogasd a munkánkat banki átutalással. Az adományokat az Átlátszónet Alapítvány számlájára utalhatod. Az utalás közleményébe írd: „Adomány”, köszönjük!