
Biotechnológikaland
Ukrajna és a GMO – Dakotarege
(Géntechnologikaland No78) Ukrajnában a GMO-k társadalmi megítélése összességében óvatos, sokszor elutasító. A közvélemény-kutatások és piaci tapasztalatok szerint a lakosság jelentős...


(Biotechnológikaland No42)
Mégis csak van mit javítani a tökéletes módszereken? A kutatás – ez az alaptermészete – ismételten sok pontosítással lepte meg a világot. Közülük az egyik nagy ígéret a genomszerkesztés. A transzgenikus eljárásokat is már hangoskodóan éltetők hirtelen pálcát törtek a termékeket eredményező korábbi módszerek felett, már ők is sorolják azok gyöngeségeit. Teátrálisan vonalat húznak: ez volt eddig, és ez következik ezután. Poétikusan a genomszerkesztés hajnaláról beszélnek, amelyben egy második Nap is felkel. Új istenek érkeztek az Olimposzra. Régi/új helytartóik most lerombolják az eddig épült templomokat. Vajon ők lennének az utolsó érkezők, vagy az igaziak majd ezután jönnek, hiszen a mostaniak már legalább meg tudják fogalmazni a géntechnológia felnőttkori álmait. Gyakorlati eredményük még kevés, de az eljárási szabadalmak már olvashatók. Nem ez lenne az, ami miatt Európa eddig is annyira tartózkodó volt?

A mára súlyosan elavult/hiányos hazai géntörvényből érthetően hiányzik két évtized géntechnológiai tudása, és az állat-géntechnológia átgondolása. A mai szabályozás még a mezőgazdasági géntechnológiára sem képes kiterjedni, nemhogy a géntechnológia egészének áttekintő szabályozására, amire szükség lenne. Az 1998-ban létrehozott hazai géntörvény géntechnológiai tartalma évtizedeken keresztül nem változott. Eredetileg kerettörvénynek készült, de általános irányelvei/útmutatásai már meghaladottak. Géntechnológiai tartalmában, rendeleti kapcsolatrendszerében szakszerűen nem egészült ki. A húsz éves elmaradás sürgős intézkedést igényel. A feladat egyáltalán nem egyszerű, az elvégzendő munka talán a mai köztisztviselők rálátási képességét meghaladja. Feltételezem, hogy ezért nem tudtak eddig lépni. A törvénykezési tétlenség azzal jár majd, hogy a törvény alá nem tartozó fejlesztési formák (az Egyesült Államokban már nem tartozik GMO körbe több laboratóriumban géncsendesített fajtacsoport) kimaradnak a jogi szabályozás alól. Ezt akarjuk valójában? Nézzük, akkor a növényekre vonatkozó részleteket; miről szólhat a jelenleg futó Genomszerkesztési Road Show, amely mintha visszavágója lenne a mezőgazdasági tárca által öt évvel ezelőtt szervezett dzsemborinak.
Helyspecifikus nukleáz-bázisú genomszerkesztés
A genomszerkesztés szóra fordítsunk egy csöpp figyelmet. Megszokhattuk már a hazai géntechnológia extrovertált és eufemisztikus szóhasználatait. Génsebészet, mondja a hazai szakértő a vaktában örökítőanyagba lőtt partikulumokra. Igazán vicces. Géntechnológiával nemesített, mondja az iparos, aki hagyományos módszerekkel kiválogatja a sok korlátozottan életképes mutáns közül a rendben lévőt. Mire jó a szépelgés? Vágyainkat nevezzük még meg ezekkel a terminusokkal. Mit fogunk, majd akkor mondani, ha megérkezünk, amikor indokolt lehet a használat. A genomszerkesztés már megfelel a génsebészet kritériumának?
Növényeknél viszont dehogy is szerkesztjük még a genomot. Ezt talán a xenotranszplantáció kapcsán mondhatja el magáról az a csoport, amely a humanizált sertésen dolgozik. Növényeknél a legjobbak egy-egy helyen (valóban csak?) ügyesen és értőn belenyúlnak az örökítőanyagba. A statisztikái ezeknek az új módszereknek sem túlságosan jók még, de ma ez is derekas eredmény. Átmenetileg. Talán ez az út vezet a tartalmi génsebészet felé, ha javítani is tudunk majd a módszerekkel. Elvileg nem reménytelen, bár ma még inkább elrontunk vele valamit, szép nevet adva ennek: csendesítünk. Értem én a döntéshozóknak szóló nyelvet, ahol sokan úgy gondolják, hogy kell a nagyot mondás. Én ezt nem szoktam díjazni. Az alul magyarázással kialakuló szellemi fölény gyakorlása is szerintem kontraszelektív. Felháborítja a fogyasztót. Fel kellene végre ébredni: egyszerűség és őszinteség, amit a tudománytól az emberek várnak.
– Meganukleázok (MN)
A 2012-es felsorolás nem említi a baktériumokból már akkor széleskörűen ismert meganukleázok használatát, bár ma már nyilvántartott módszer a genomszerkesztés területén (lásd LAGLIDADG homing meganuclease – 49. ábra). A további nukleáz-bázisú módszereket a természetes meganukleázok limitált használhatósága miatt fejlesztették ki. A hosszú felismerő szakasz miatt ezt tartják a legpontosabb módszernek. Mára módosított ún. hibrid meganukleázok használatára épülnek az E-Drel és DmoCre módszerek. A tervezett meganukleázok szabadalmi védettsége 2005-ben indult. Kombinált módszer a megaTALs, amely a TALENs kombinált felhasználásával jött létre. Az E-Drel segítségével a Bayer kutatói pontosan megtervezett helyű (GHB119) módosítást hajtottak végre a gyapotban, ahová a hppdPF (isoxaflutol-, tembotrione-tűrés) és a 2m epsps (glyphosate-tűrés) géneket helyezték el, a korábban bevitt cry2Ae (hernyó-rezisztencia) és bar (glufosinate-tűrő) transzgének mellé. Az átalakítás sikeressége 2% volt, ami mindjárt el is gondolkodtat. A módszer az egyszeresen módosított fajtacsoportok gyors többszörössé módosítására nyitott lehetőséget. Ami ennél is fontosabb, hogy megcélzott helyre (locus) való beépítésre is van lehetőség, ami a szűkebb értelemben vett (sensu stricto) ciszgenikus növények irányába vezethet.
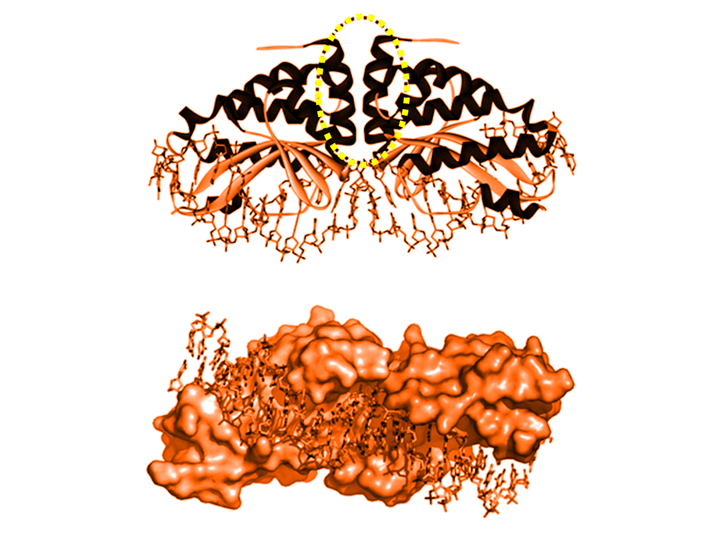
49.ábra: Módosított, I-Crel meganukleáz és a cél-DNS. Felső képen a LAGLIDADG motívum sárga/barna szaggatott oválissal jelölve; az alsó képen a DNS-spirál illeszkedése (Arnoud et al./PEDS nyomán)
– Cinkujj-nukleázok
A cinkujj-nukleázok (zinc-finger nuclease, ZFN) cinktartalmú enzimek, amelyek ún. cinkujj fehérjedoméneket (cinkion által stabilizált alegységeket) tartalmaznak, melyek egyenként az enzim által hasításra kerülő nukleinsavban egy-egy adott triplet felismerésére képesek. Az itt felsorolt módszerek közül ez a legcitotoxikusabb, ugyanakkor a használhatósága nagyságrendekkel jobb, mint a természetes meganukleázoké. A nukleázok a DNS mindkét, míg a nikázok az egyik szál bontására képes enzimek. A természetes cinkujj-fehérjékben 3-4 cinkujj kapcsolódik egymáshoz (50. ábra). Szintetikusan előállítható 6-12 egységet tartalmazó konstrukció is, amely 18-36 bázisnyi DNS-szakaszt ismer fel és kötődik ahhoz. Azt gondolhatjuk, hogy minél hosszabb az azonosító szakasz annál pontosabb a cél felismerése, így annál kisebb esélyű a nem kívánatos helyen való hasítás. Ez azonban a ZFN módszernél nincs így. A hosszabb felismerési szakasz itt több helyet ismerhet fel közel azonosnak, és ezáltal hasíthat olyan helyen, ahol nem kellene. A DNS-szál törése tipikus mutációs esemény, amelyet a sejt DNS-javító mechanizmusa igyekszik kezelni. A ZFN-konstrukcióhoz a FokI hasító enzim kapcsolódik, amely a felismert helyen vágja a célzott DNS-t, és mindkét szál törését okozza. A ZFN-fehérjéket azonban nem lehet közvetlenül a sejtekbe bevinni (lásd Venetianer), viszont az ezeket kódoló DNS-t vagy mRNS-t igen, vagyis speciális plazmid készíthető ilyen céllal.
A vektorplazmid DNS-e által kódolt endonukleáz megtalálja a genomban a megfelelő részt, ott kapcsolódik ahhoz, és elvágja a célzott DNS mindkét szálát. A sejtben ekkor a DNS-törés kezelésére javító folyamat indul el, ami mintaszekvencia nélkül (nonhomologous end joining, NHEJ) gyakran hibás végeredményű, aminek következtében a gén megsérülhet és funkció képtelenné válik (vö. csendesítő génmutáció). A ZFN három változata ismert. ZFN-1 esetében a cinkujj-nukleázt kódoló transzgén nem tartalmaz javító templátot. A ZFN-2 néhány kilóbázis hosszúságú és a célzott génszakasszal homológ javító templátot (vö. homology-directed repair, HDR) tartalmaz. Ekkor tervezhető, hogy miként rontsa el a nem kívánatos gént. A ZFN-3 a végein a hasítási hely két oldalán lévő DNS-szekvenciákkal homológ, akár több kilóbázis hosszúságú DNS-szakaszt tartalmaz. Funkcionális értelemben a ZFN-3 hosszú DNS-szakaszok (transzgének és allélek) célzott beépítésére (transzgenezis és ciszgenezis), míg a ZFN-1 és ZFN-2 pontmutációk megvalósítására (vö. mutációs alapú géncsendesítés) alkalmasak.
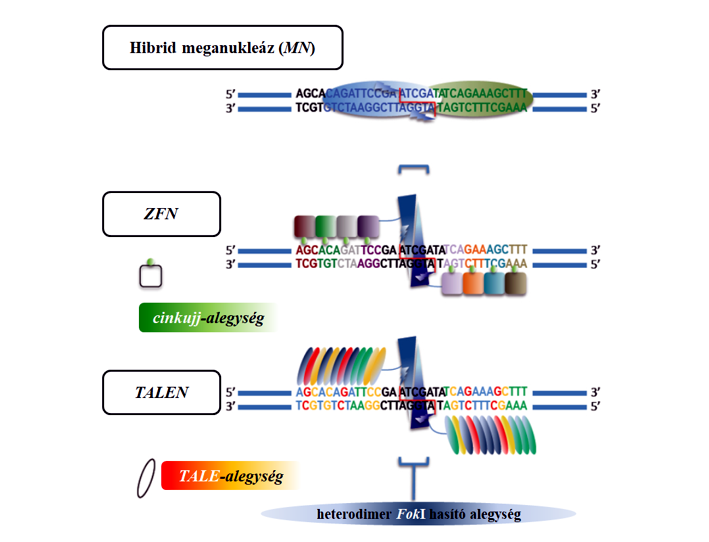
50.ábra: Genomszerkesztési eljárások: hibrid meganukleáz, ZFN és TALEN (Farzad Jamshidi nyomán)
A ZFN módszernél tehát egy erre a célra tervezett plazmidon lévő génkazettában genetikai információ kerül a sejtbe, amelyről a sejtben fehérje íródik át. A hasítás nem feltétlenül pontos, vagyis hasonló, nem tervezett helyeken is megvalósulhat, ami akár a sejt pusztulásához is vezethet. Ez az oka annak, hogy nem terjedt el a humán-egészségügy gyakorlatában, ahol nincs tervezhető vesztesség. A donorsejtbe bekerülő, a változást végrehajtó teljes genetikai információ ugyan nem épül be az örökítő rendszerbe, de pontmutációt okoz (ZFN-1 és ZFN-2), úgy hogy elront egy gént, ami viszont nem feltétlenül hallgat el, bár a korábbi funkcionális fehérjét nem produkálja. Ha nem annyira sérült (a saját reparációs rendszer állítja helyre a működést), akkor termelhet valamilyen fehérjét (ZFN-1) aminek a sorsa mindenképpen feltárásra szorul. A javító templátot tartalmazó változat (ZFN-2) ennél jobb megoldás, hiszen az elrontás mikéntje is kódolt. Beépíthet a rendszer (ZFN-3) olyan tervezett allélt/gént/DNS-szakaszt is, amely a genom egy tulajdonságát változtatja meg (51. ábra).
2009-ben publikálták azt az eredményt, hogy lúdfűbe (Arabidopsis thaliana) dohány acetolaktáz-szintetáz géneket (als SuRA és als SuRB) ZFN-3 módszerrel vittek be, amelyek imidazolinone és szulfonilkarbamid-tűrést eredményeztek. Hasonló módon állítottak elő és szabadalmaztatták a Dow és Sangamo kutatói transzgenikus, glufosinate-tűrő (pat) kukoricát 2009-ben. A pat gént az ipk1 gén helyén vitték be, amit így működésképtelenné tettek. Ennek következményeként a transzformált növények nem csak glufosinate-tűrők lettek, hanem a foszfor-tárolására használt fitát termelődése lecsökkent, ami kedvezőbb takarmányt, így környezetbarát trágyát (vö. foszforterhelés) eredményezett. Ezzel a módszerrel allél bevitel (m epsps, 2m epsps), így ciszgenikus glyphosate-tűrő fajta is előállítható, bár ez nagyságrendekkel komolyabb feladat, mint a transzgenikus fajtaváltozat elkészítése.
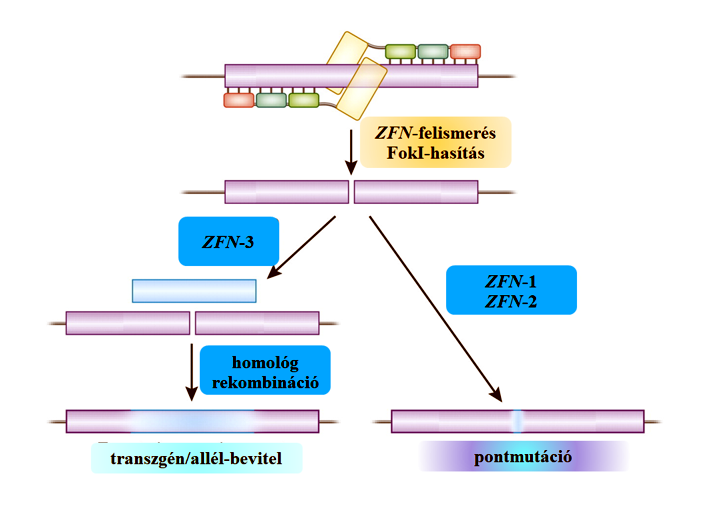
51.ábra: Cinkujj-nukleázokkal (ZFN) való genomszerkesztés (Carroll és Wahl, 2011/GSA nyomán)
Az átalakításhoz használt plazmidról/génkazettáról is ejtsünk pár szót (52. ábra), hiszen annak sorsa rendkívül kritikus, ha a korábbi transzformálásokkal való azonosságot vagy különbözőséget keressük. Nos, úgy tűnik, hogy ezen a szinten több a megegyezés, mint az eltérés a korábbi megoldásokkal. A kétségkívül fajidegen elemeket tartalmazó plazmid sorsát illetően – szabadalmi okokból – a leírók nagyon szűkszavúak. Lehet ezt a sejtrutinra bízni (gyorsan egységeire bomlik – mondják), de az engedélyezés szintjén a hitre támaszkodni talán mégis kevés. Ember esetén a kis arányú céltévesztés sem megengedhető. Térjünk vissza a ZFN-módszer (a többi itt felsorolt módszerre is kiterjeszthető ez a tudás) specifikusságára, amelyre vonatkozóan súlyos kritikai eredmények jelentek meg tudományos lapokban, ami szerint a kiválasztott hasító helynek megfelelő/hasonló a genomban sokkal több létezik, mint amire a rutinvizsgálataink általában fókuszálnak. Ezek az átalakítások ugyancsak megtörténnek (az egész genomra kiterjedő off target hatásokat kellene vizsgálni), és esetleg észre sem vesszük ezeket. Mindez még jelentősebb arányban fordult elő a CRISPR/Cas9 endonukleáz módszer használatakor. Ezek a felismerések további módszerfejlesztésre intenek (valójában nem érkeztünk még meg, csak úton vagyunk), és az egészségügy ezért nem engedte még be ezeket a gyógyítás eszközei közé.
A jelenleg felhasználható genetikai elemek az eddigi géntechnológiai tudásunk eredményei. Közöttük nem látható még az azonos fajból származó ’promóter–intron–allél–terminátor’ konstrukció. Mindez természetesen bekövetkezhet, de a gyakorlat számára pillanatnyilag csak ígéret a szűkebb értelemben vett ciszgenikus növényt eredményező genomszerkesztés (lásd később). A fajtacsoportokat bemutató adatbázisokban nem találunk még, így készült, kereskedelmi forgalomba kerülteket. A lehetséges megjelenésükre azonban az európai törvénykezésnek most kell felkészülni.
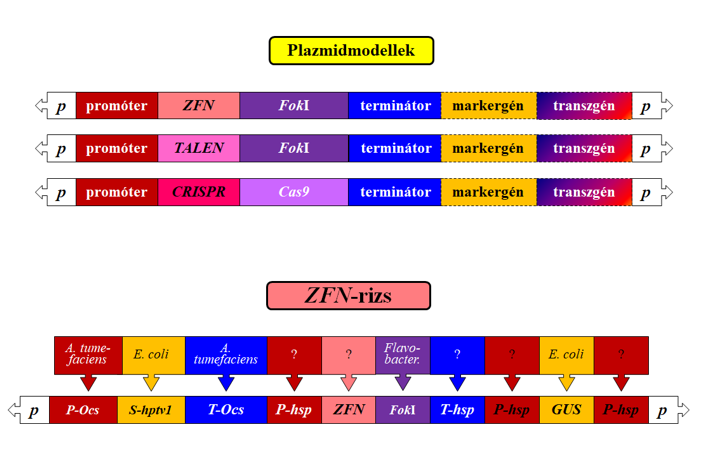
52.ábra: Plazmidvázlatok genomszerkesztéshez
– TALENs
A helyspecifikus nukleáz-bázisú genomszerkesztéshez, ahová a ZFN is tartozik, ma felsorakozik a TALENs és CRISPR/Cas9 is. A TALENs (transcription activator-like effector nucleases) nevű technika sok tekintetben hasonlít a ZFN módszerhez. Ezt Xanthomonas fajból írták le, amellyel a növénypatogén baktérium felismer a gazdában bizonyos génszakaszokat és ahhoz kötődik. A TALE alegységek egy-egy nukleotidot képesek felismerni (vagyis nem tripletet mint a ZFN, ami miatt ezt hajlékonyabb technikának tartják), és egymás mögé rendezésük hosszabb DNS-szakasz felismerésére is képes lehet (50. ábra). A FokI hasító enzimhez ez is kapcsolható és a továbbiakban a használhatósága a ZFN-nél leírtakhoz hasonló. Vannak, akik a ZFN-nél pontosabb módszernek tartják. Segítségével kísérleti körülmények között Xanthomonas oryzae pathovar. oryzae-tűrő rizsát állítottak elő, azonban úgy, hogy előtte Agrobacterium-vektort is alkalmaztak módosításra. Az Agrobacterium-vektorral bevitt transzgén a későbbi szelekció során távolították el, vagyis kombinált nemesítési eljárást alkalmaztak. A klasszikus nemesítési eljárásokkal való kombináció (pl. szelekció, visszakeresztezés) egyébként itt is a fajtacsoport- és a fajta-előállítás része.
A szójaolaj megváltoztatására a klasszikus Agrobacterium-vektoros átviteli technológiával már fajtacsoportot értek el. A MON-87705-6 (Vistive Gold) genetikai esemény a fatb1-A és fad2-1A géneket tartalmaz, amelyek következtében az olajban nő az egyszeresen telített olajsavak aránya. A TALENs módszerével fad2-1A és fad2-1B géneket vittek be, ami az egyszeresen telítetlen zsír- és olajsavakból többszörösen telítetlen zsír- és linolénsavakat eredményezett.
– CRISPR/Cas9
2012-től a fentieknél egyszerűbben kezelhető és gyorsabb technológia, a CRISPR/Cas9 (clustered regularly interspersed palidromic repeats) módszer már jelentős karriert futott be. Ha egy baktériumot első ízben ér fágtámadás, akkor az idegen DNS egy darabja beépül a baktérium DNS-ének CRISPR régiójába. Második támadás esetén, ez alapján a baktérium felismeri a támadót és elbontja. A konstrukció felismerést végző része ma a Streptococcus pyogenes baktériumból előállított gRNS (guide RNS), amely egyik fele a cél DNS megváltoztatandó szakaszával komplementer, míg a másik fele komplexet képez a Cas9 enzimmel. A gRNS készen vásárolható, és a specifitásért felelős. Az előzetes fehérjekémiai munka elmaradása így gyorssá és olcsóvá teszi a munkát. Egyidejűleg pontatlanabbá is, hiszen a célgén ismert változatával tervezünk. A DNS-regeneráció hasonló, mint korábban említettem. Ebben az esetben is bekerül a módosítandó szervezetbe idegen genetikai tartalom (a viaszos kukoricánál a Wx1-gén csendesítésére például hatféle plazmid is – lásd később), de olyan nyomot hagy, amilyet tervezünk a számára. Amennyiben allélt viszünk be, akkor ciszgenikus fajtát hozunk létre, ha a célunk géncsendesítés (mint az eddig sikeres fajtáknál), akkor az történhet, azonban transzgén bevitelére is alkalmas, vagyis elhatározás kérdése, hogy milyen szervezetet hozunk létre ezzel a technológiával. Ezért sem lenne helyes az alkalmazott módszer alapján való engedélyezési eltérés.
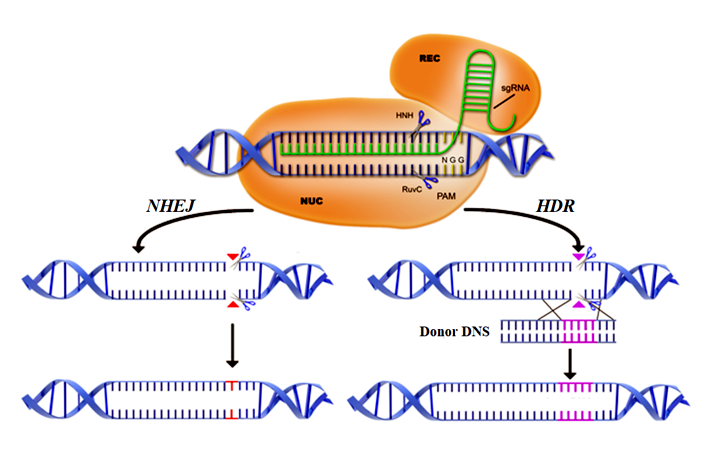
53.ábra: A CRISPR/Cas9 módszer vázlata – Rövidítések: Donor DNS – mintaszekvencia; HDR (homology directed repair) – homológ végillesztés; HNH – a cél-DNS-t hasító egység; NHEJ (non-homologous end joining) – nem-homológ végillesztés; NUC (nuclease lobe) – nukleáz lóbusz; PAM (protospacer adjacent motif) – 3-5 bázispáros szakasz, ami mellett a Cas9 hasítani képes; REC (recognition lobe) – felismerő lóbus; RuvC – a nem-cél DNS-t hasító egység; sgRNA (single-guide RNA) – egyszálas vezető RNS (Colin Gillespie nyomán)
A helyspecifikus nukleáz-bázisú genomszerkesztési módoknak hátrányai is vannak, amennyiben kis valószínűséggel más, nem tervezett helyeken is vághat a konstrukció, s ez új mutációkat eredményezhet. A CRISPR/Cas9 ebből a szempontból a felsoroltak közül a leggyengébben teljesítő változat. Gyors, olcsó (emiatt egyesek lehetséges biohacker-ekről elmélkednek, akik a nyári konyhából szabadítanak majd a világra eddig sohasem létezett szervezeteket – a gene drive kihasználására gondolni sem akarok), de még pontatlan, vagyis a vele kapcsolatos lelkesedés talán még korai. Javítására már vannak elképzelések (CRISPR/cpf1, CRISPR/Cas9D10A nikáze stb.). Messze még az út vége, és nem születtek még meg azok a döntéshozók sem, akik kezében mindez az emberiség számára hasznos eszköz lehet.
A következő rész címe: Genomszerkesztés 4 – Átalakítjuk a bolygónk élővilágát? (Biotechnológikaland No43)
Darvas Béla
Bankszámlaszám: 12011265-01425189-00100001
Bank neve: Raiffeisen Bank
Számlatulajdonos: Átlátszónet Alapítvány
1084 Budapest, Déri Miksa utca 10.
IBAN (EUR): HU36120112650142518900400002
IBAN (USD): HU36120112650142518900500009
SWIFT: UBRTHUHB
Számlatulajdonos: Átlátszónet Alapítvány
1084 Budapest, Déri Miksa utca 10.
Bank neve és címe: Raiffeisen Bank
(H-1133 Budapest, Váci út 116-118.)
Támogasd a munkánkat az Átlátszónet Alapítványnak küldött PayPal-adománnyal! Köszönjük.
Támogatom PayPal-adománnyalHa az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
1% TÁMOGATÁS


Támogasd a munkánkat 10 ezer forint adománnyal, mi pedig megajándékozunk egy pólóval. Katt a részletekért.
Támogasd a munkánkat palackvisszaváltással, kattints az üvegvisszaváltós oldalra, mentsd el a kódunkat, és használd azt a Repontoknál!
Ha van bankkártyád, akkor pár kattintással gyorsan tudsz rendszeres vagy egyszeri támogatást beállítani nekünk az adjukossze.hu oldalán.


Postai befizetéssel is tudsz minket támogatni, amihez „sárga csekket” küldünk. Add meg a postacímedet, és már repül is a csekk.
Néző, Szurkoló, B-közép és VIP-páholy kategóriás Átlátszó-előfizetések között válogathatsz a Patreonon.
Bárhol is dolgozol a világban, ha a munkáltatód lehetőséget ad arra, hogy adott összeget felajánlj egy nonprofit szervezetnek, akkor ne feledd, a Benevity-n keresztül az Átlátszónet Alapítvány is ajánlható.
Ha az 1 százalékodat az Átlátszó céljaira, projektjeire kívánod felajánlani, a személyi jövedelemadó bevallásodban az Átlátszónet Alapítvány adószámát tüntesd fel: 18516641-1-42
(Géntechnologikaland No78) Ukrajnában a GMO-k társadalmi megítélése összességében óvatos, sokszor elutasító. A közvélemény-kutatások és piaci tapasztalatok szerint a lakosság jelentős...

(Biotechnologikaland No77) A Marton Genetics Kft. állásfoglalása A Marton Genetics (Bázismag Kft.) cégcsoport a martonvásári kutatóintézet (Agrártudományi Kutatóközpont) munkájára épült....
(Biotechnologikaland No76) Előszó A GMO-lobbi hatására az Európai Bizottság javaslata (címábra) komoly visszhangra talált Európában. A javaslatok közül NGT1 esetében...

(Biotechnologikaland No75) Barabás Zoltán Biotechnológiai Egyesület kontra GMO-Kerekasztal A Barabás Zoltán Biotechnológiai Egyesület lépett hazánkban először abból a célból, hogy...
Támogasd a munkánkat banki átutalással. Az adományokat az Átlátszónet Alapítvány számlájára utalhatod. Az utalás közleményébe írd: „Adomány”, köszönjük!